Для установления возраста, в котором появляется реакция на потенциальную жертву в процессе постнатального онтогенеза, волчатам с 20-дневного возраста через день предъявляли молодых крыс, наблюдая за их реакцией. Параллельно определяли мотивационную сущность реакции, возникающей у волчат при предъявлении им потенциальной жертвы. Важно было определить, как они ее воспринимают как объект питания или как движущийся предмет.
Для выяснения этого вопроса использовали радиоуправляемые подвижные тележки, на которые попеременно прикрепляли деревянные бруски, чучело крысы или кролика, изготовленное из проветренной или искусственной шкуры, тушки крыс или кроликов. Тележки попеременно двигались то в направлении хищника, то от него. Во всех случаях регистрировали поведение волчат до полной потери у них интереса к предлагаемому предмету. Одновременно с радиоуправляемыми предметами волчатам предъявляли взрослых лабораторных крыс и кроликов. В этих случаях наблюдали за способностью умерщвления и поедания жертвы волчатами. Такие эксперименты проводили с интервалом от 2 до 7 дней. Параллельно изучали способность формирования хищнического поведения путем опосредованного обучения (9 индивидов), т. е. за счет наблюдения за действиями взрослых особей или сеголеток.
Таким же методом изучали принципы формирования хищнического и охотничьего поведения у кошачьих. На первом этапе экспериментов этим хищникам предъявляли взрослых кроликов, на втором (за исключением пум) — коз.
Для оценки возрастных изменений способности к обучению неопытным волкам (n=8, возраст — от 7 месяцев до года) предъявляли разнообразный домашний скот и регистрировали хищническое поведение.
При изучении принципа формирования приемов нападения на жертву у волков, имеющих опыт охоты на мелких животных (возраст-1 год, n=3; и 2 года, n=3), им предоставляли возможность охоты на ослов (18 случаев). При этом определяли как направление первой атаки, так и участки тела жертвы, которые первыми подвергались нападению.
В отдельных экспериментах изучали формирование навыков групповой охоты. Эксперименты велись на шести группах волков (I — n=3; II — n=4; III — n=6; IV — n=4; V — n=5; VI — n=4).
Члены I, IV и VI групп не имели опыта охоты на крупную жертву, среди членов II группы такой опыт имел только один волк, из III группы — два волка, из V — три. Каждой группе жертва предъявлялась 4 раза. В каждом конкретном случае регистрировали взаимодействие каждого хищника с жертвой и друг с другом.
При изучении постнатального онтогенеза поведения обращали внимание на формирование приемов игры хищных между собой. Особый интерес для нас представляло значение игры для формирования приемов охоты.
В процессе наблюдения регистрировали все приемы, которые молодые животные применяли во время игры друг с другом и с неживыми предметами, и сравнивали их с приемами, применяемыми во время охоты как дикими, так и выращенными в неволе взрослыми волками.
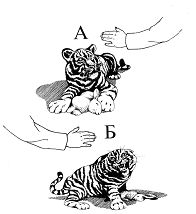
Эксперименты показали, что у волчат месячного возраста, независимо от пола, полностью сформированы реакции преследования и схватывания потенциальной жертвы. Эти реакции сформированы также у находившихся в эксперименте трехмесячных кошачьих. В случае поимки жертвы хищники периодически встряхивали ее до тех пор, пока она не переставала двигаться. После этого они, как правило, бросали жертву и начинали интенсивно ее обнюхивать. Параллельно с этим у них время от времени возникала реакция «пробы на зуб» (рис. 10А). При этом хищники часто перемещали жертву лапами, а если она двигалась или перемещалась самостоятельно начинали играть с ней (рис. 10Б).
Рисунок 10.
Реакция неопытного волчонка на потенциальную жертву (рисунок В.М. Смирина).
А — исследование с «пробой на зуб; Б — игровое поведение.
Поедание жертвы не наблюдалось ни в одном из случаев. Описанные реакции оставались типичными для представителей обоих семейств.
У 40—50-дневных волчат при виде удаляющихся предметов возникала реакция преследования (рис. 11А) и схватывания. При непосредственном контакте с ними у животного развивалось игровое поведение. Эти реакции полностью формируются к 2—3-месячному возрасту. При развитии игрового поведения хорошо выражена реакция «пробы на зуб». В случае остановки предмета волчата некоторое время продолжали играть и исследовать предмет, при этом часто перемещая его лапами. В конечном счете животное полностью теряло интерес к предмету. Принципиально схожие реакции развивались и у всех находящихся в эксперименте кошачьих.
Рисунок 11.
Реакция на направление движения предмета.
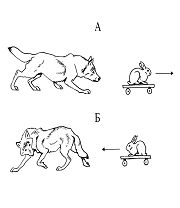
А — преследование предмета, движущегося в сторону от хищника;
Б — избегание предмета, движущегося в сторону хищника.
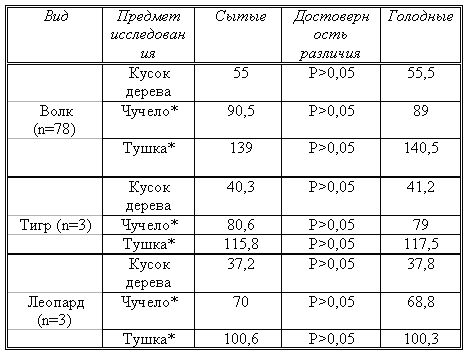
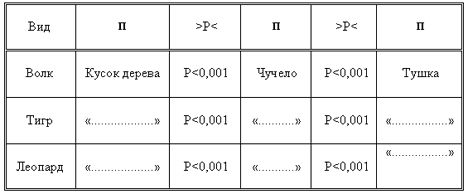
Во всех случаях мы регистрировали время, в течение которого животные исследовали попадающие в поле их зрения предметы. Это время определялось как для голодных, так и для сытых животных. Статистический анализ материала показал, что время, затраченное голодными и сытыми животными на исследование предметов одинакового происхождения, не различалось (во всех случаях р › 0,05). Из таблицы видно, что изменяется продолжительность времени при исследовании предметов различного происхождения, где p ‹ 0,001 (таблица № 5а, б).
В тех случаях, когда предметы двигались в сторону хищника, у него появлялась четко выраженная реакция избегания (рис. 11Б), а в некоторых случаях он отскакивал в сторону. Последняя реакция лучше была выражена у кошачьих. После 2—3 таких взаимодействий животные целенаправленно отскакивали в сторону таким образом, чтобы напасть на предмет сзади.
Таблица 5a.
Средняя продолжительность исследования предметов разного происхождения. * — кролик.
Таблица 5б.
Достоверность различия времени исследования предметов разного происхождения. П — предмет исследования.
У хищников того же возраста при предъявлении им живой потенциальной жертвы (крысы, кролики) развивались такие же реакции, как и на радиоуправляемые предметы. Хищники убивали потенциальную жертву, как правило, при ее попытке к бегству. Это происходило из-за сильного смыкания челюстей при схватывании. После умерщвления жертвы (в случае, если шкура не повреждалась), продолжалась игровая и исследовательская активность с «пробой на зуб». В процессе исследования особый интерес животных вызывали область живота и уши жертвы.
Через некоторое время животные утрачивали интерес к обездвиженной жертве и покидали ее. В тех случаях (n=38 для волков и n=2 для кошачьих), когда у жертвы при поимке повреждалась шкура, хищники прекращали всякую активность, кроме обнюхивания и облизывания поврежденного места, после чего съедали жертву. При следующем предъявлении потенциальной жертвы они демонстрировали полностью сформированное хищническое поведение, без всякой игровой и исследовательской активности. Животные преследовали жертву, ловили ее и съедали.
Рисунок 12.
Реакция неопытного в хищничестве зверя на потенциального конкурента.
А — у жертвы шкура не повреждена; Б — у жертвы шкура повреждена.
Особо следует подчеркнуть, что сильно различались реакции на потенциального конкурента в случае повреждения шкуры жертвы и тогда, когда она оставалась неповрежденной. Если шкура оставалась неповрежденной, при появлении потенциального конкурента (сородича или экспериментатора) хищники не реагировали на него, а в случае повреждения шкуры жертвы у них наблюдалась ярко выраженная агрессия по отношению к конкуренту (рис. 12 А, Б).
Исследуя хищническое поведение у животных разного возраста мы выяснили, что у неопытных хищников (n=7, от 7 месяцев до одного года) при виде потенциальной жертвы развивалась только исследовательская активность, без включения игровых элементов. Во время приближения потенциальной жертвы, независимо от ее скорости, у хищников развивалась пассивно-оборонительная реакция (рис. 13 А, Б, В, Г), а при увеличении дистанции между ними — хорошо выраженная реакция преследования и схватывания. В тех же случаях, когда жертва оказывала сопротивление, у хищных развивалась типичная агрессивная реакция со всеми присущими ей компонентами.
Рисунок 13.
Последовательность реакций неопытного годовалого волка на потенциальную жертву.
А — дистантное исследование;
Б — исследование при начальной стадии избегания;
В — начальная стадия пассивно-оборонительной реакции;
Г — рычание, во время пассивно-оборонительной реакции.
Волчата этого возраста съедали жертву только в том случае, если у нее повреждалась шкура. В тех случаях (n=4), когда шкура оставалась неповрежденной, после кратковременной исследовательской активности волки теряли к жертве всякий интерес, съедали ее только тогда, когда экспериментатор сам повреждал ее шкуру. При последующем предъявлении волкам потенциальной жертвы у них развивалось полноценное хищническое поведение.
По-другому развивались события, когда пятерым 2-летним волкам, не имеющим опыта хищничества, впервые предъявляли потенциальную жертву (овцу). Во всех случаях (10 опытов с каждым волком) у животных появлялась только дистантная исследовательская активность. Расстояние между ними сокращалось не больше чем на 2 м. Когда потенциальная жертва приближалась к хищнику, он отступал или отскакивал, сохраняя при этом «безопасную» дистанцию. Также развивались события и после предоставления хищникам возможности наблюдения за охотой опытных сородичей. Попытка охоты проявлялась только после того, как особям хотя бы два раза давали заранее убитую, не освежеванную жертву.
При исследовании принципов формирования приемов единоборства и обучения во время охоты на крупную жертву мы обнаружили, что хищник, как правило, целенаправленно обходил жертву и нападал на нее сзади. Первое нападение приходилось на круп, реже — на хвост или задние ноги. В тех случаях, когда попытка покорения жертвы оказывалась безуспешной, волки атаковали разные части ее тела. При последующей охоте хищники во время нападения на жертву использовали прием, оказавшийся успешным при предыдущей охоте.
Исследования принципов формирования навыков групповой охоты волков показали, что члены группы, не имеющие опыта охоты на крупную добычу, нападают на нее сзади и хватают за круп, хвост или задние ноги. При первой атаке хищники мешали друг другу, так как расстояние между ними было слишком маленьким. Уже после нескольких первых атак это расстояние увеличивалось до оптимального, после чего животные больше не мешали друг другу. Следует отметить, что, когда расстояние между особями увеличивалось, возрастало и количество атакуемых участков тела жертвы, в результате чего покорить ее было гораздо легче. В процессе апробирования разнообразных вариантов расстояние между индивидами оставалось оптимальным.
Наблюдения за остальными группами показали, что и в них события развивались аналогично описанным. Разница состояла лишь в том, что опытные члены группы начинали охоту с приема, оказавшегося успешным при предыдущей охоте, а неопытные — атаковали жертву сзади и хватали ее за круп, хвост или ноги.
Многочисленные наблюдения за каждой группой показали, что в процессе накопления опыта единоборства с крупной жертвой у хищников развивалась тенденция преимущественной атаки определенных участков тела жертвы. Такая специализация обусловлена физическими и психологическими особенностями индивидов. Так, например, слабые особи атакуют внешнюю сторону бедра или область лопаток, сильные — живот, голову или ноги. В тех случаях, когда одного из членов группы отпускали для нападения на жертву позже остальных, он вначале атаковал свободную от нападения часть тела, но затем в течение нескольких секунд перемещался к желаемой области.
Обсуждение принципов и механизмов формирования хищнического и охотничьего поведения мы проводили в соответствии с последовательностью их развития в процессе постнатального онтогенеза.
До обсуждения полученного материала следует уточнить, что до сих пор подразумевалось под врожденным хищническим поведением. Это облегчит формулирование выводов из полученного материала. Под врожденным хищническим поведением подразумевается тот врожденный поведенческий комплекс, который направлен на поимку и поедание жертвы.
Из полученных результатов следует, что реакция преследования и схватывания потенциальной жертвы у волков полностью формируется к месячному возрасту. К сожалению, нам не удалось установить время формирования этих реакций у кошачьих, так как мы получили этих животных уже в возрасте 3 месяцев, когда данные реакции у них уже были полностью сформированы. Нам достоверно известно, что до того, как они попали к нам, их кормили молодыми кроликами. Принимая во внимание результаты экспериментов с подвижными радиоуправляемыми предметами, мы пришли к выводу, что указанные реакции не являются составной частью пищедобывательного процесса. Эти реакции — определенная форма исследовательского поведения. Реакции хищников на движущиеся объекты описаны у Эйбл-Эйбесфельдта (Eibl-Eibesfeldt, 1961), Фокса (Fox, 1969), Винсента и Бэкоффа (Vinsent amp; Bekoff, 1978). По мнению этих авторов, движение потенциальной жертвы вызывает ее преследование хищником. Фокс (Fox, 1969) считает, что игровое поведение, элементы которого мы наблюдали при первом взаимодействии молодых животных с потенциальной жертвой, является организующим фактором ее умерщвления. Юэр (цитировано по Vinsent amp; Bekoff, 1978) подчеркивает, что поведение, которое в таких случаях интерпретируется как игра, возможно, является активностью, возникающей при неудаче в достижении цели. В свое время мы отмечали, (Бадридзе, 1987 а, б), что элементы игры часто появляются при сильном нервном напряжении и являются одним из инстинктивных и вместе с тем эффективных способов его снятия. В описанном нами материале возникновение этой формы поведения станет понятным, если учесть, что при исследовании неизвестных объектов у животных появляется сильное нервное напряжение (т. н. «эффект новизны»). На фоне описанного поведения возникновение игры является той инстинктивной формой поведения (смещенная активность), которая облегчает непосредственный контакт хищника с потенциальной жертвой при первой встрече.
Как видно из нашего материала, умерщвление потенциальной жертвы при первом контакте с ней не совершается целенаправленно. Если она оказывает сопротивление во время ее «пробы на зуб», у хищника развивается агрессивная реакция, что увеличивает вероятность случайного умерщвления жертвы.
Анализ исследования хищниками различных движущихся объектов показал, что наибольший интерес у них вызывают предметы, обладающие животным запахом и покрытые шерстью. Наличие такой реакции несравненно увеличивает вероятность непосредственного контакта с жертвой и в конечном счете приводит к ее умерщвлению. Как показал Фентресс (Fentress, 1967), множество псовых преследуют тот объект, который удаляется от них и, в то же время, убегают от движущегося на них объекта. В наших экспериментах схожие результаты получены на представителях обоих семейств. Однако следует подчеркнуть, что и псовые, и кошачьи очень скоро обучаются целенаправленно пропускать движущийся на них предмет и нападать на него сзади. Способность к такому обучению делает в дальнейшем более безопасной охоту на крупную жертву, фронтальное столкновение с которой может быть опасным для хищника. Согласно нашим данным, для формирования хищнического поведения необходимо чтобы животное убедилось в съедобности потенциальной жертвы. Осуществление же самого акта еды происходит после восприятия запаха, вкуса крови и ее вида. Это подтверждается и тем фактом, что хищники не охраняют от потенциальных конкурентов убитую ими добычу, если ее шкура не повреждена. При повреждении же шкуры жертвы ее защита происходит на фоне ярко выраженной агрессивной реакции. Именно после первого поедания жертвы у хищных формируется хищническая реакция, как единый поведенческий комплекс.
Из вышесказанного можно заключить, что реакция неопытного хищника на потенциальную жертву является составной частью исследовательского поведения, а хищническое поведение, как форма пищевого поведения развивается после установления хищником факта ее съедобности. Мотивационная основа первого и второго абсолютно различны. В свое время Винсент и Бэкофф (Vinsent amp; Bekoff, 1978) высказали мнение, что, поскольку голод и питание связаны друг с другом, то с ними должно быть связано и хищническое поведение; они допускают, что койоты, возможно, «ассоциируют» жертву с едой. Согласно нашим данным, подобная ассоциация возникает именно после установления факта съедобности жертвы. У неопытных хищных питание и умерщвление потенциальной жертвы мотивационно не связаны друг с другом. Исходя из этого, можно заключить, что хищническое поведение представляет собой форму пищевого поведения, мотивация которого возникает при восприятии живого, потенциального объекта питания. Охотничье же поведение подразумевает форму пищедобывательного поведения, которое может развиться только после формирования хищнической реакции и на ее базе.
Итак, реакция неопытных и опытных волчат на потенциальную жертву различается по своей мотивационной основе и является в первом случае — исследовательской, а во втором — пищедобывательной (охотничьей). Врожденное предпочтение, которое неопытные животные отдают предметам, покрытым шерстью и имеющим животный запах, повышает вероятность их контакта с потенциальной жертвой со всеми вытекающими из этого последствиями.
Наблюдения показали, что у молодых животных хорошо выражена способность обучения хищничеству, опосредованного наблюдением за другими особями. Как отмечалось в главе 2.1, такой же феномен имеет место и у дикоживущих молодых волков. Существование его значительно упрощает и, в конечном счете, ускоряет формирование хищнического поведения. Это становится особенно важным, если учесть, что у 7-месячных волков, не имеющих опыта охоты, значительно затруднен процесс непосредственного контакта с потенциальной жертвой, а с 2-летнего возраста это практически невозможно. По нашему мнению, основной причиной этого является исчезновение некоторых врожденных реакций, проявляющихся только в определенные периоды постнатального онтогенеза в процессе постнатального онтогенеза. В этом случае игра выполняет функцию снятия нервного напряжения.
При изучении принципов формирования поведения охоты на крупную жертву мы обнаружили, что у хищных ярко выражена реакция нападения на жертву сзади. Эту реакцию, на первый взгляд, следует отнести к разряду инстинктивных реакций, так как, согласно существующим в литературе данным (Mech, 1975; Hanshaw et al, 1979; Leyhasen, 1979) и нашим наблюдениям, этой реакцией обладают как неопытные, так и имеющие опыт хищные. Для полного освещения данного вопроса необходимо привести наш материал о развитии игрового поведения между сеголетками.
В данном случае игра представляет интерес с точки зрения ее так называемой экзогенной функции. Исходя из этого, игра с партнерами будет рассматриваться соответственно с позиции «теории практики», автором которой является Карл Гросс (Groos, 1898). Эту теорию в дальнейшем развили Алдис (Aldis, 1975) и Ллойд-Морган (по Фабри, 1993). Рассмотрение полученного нами материала с этой позиции дает возможность определить значение игры как в процессе формирования приемов единоборства с крупной жертвой, так и охотничьего поведения взрослых животных в целом.
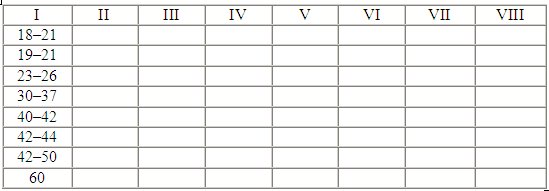
Полученный материал свидетельствует, что у волчат игровые элементы впервые появляются в возрасте 18—21 дня, что близко литературным данным (Фабри, 1993). Понятно, что появление игровых элементов сопровождается увеличением интенсивности моторной активности. Следует отметить характерные для этого периода следующие виды моторной активности: покусывания в горло или в шею, удары лапами. Этапы формирования игровых элементов приведены в таблице 6. Как видно из таблицы, появление новых элементов игры завершается к 2-месячному возрасту. Усложнение игры происходит в последующий период за счет совершенствования отдельных элементов и увеличения количества вариантов их комбинирования.
Таблица 6.
Этапы формирования разных элементов игрового поведения.
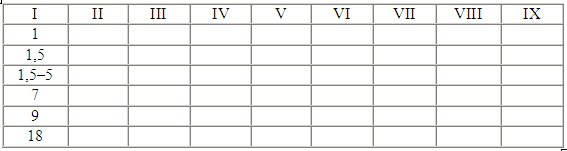
I — возраст в днях;
II — удар лапами;
III — хватка за горло или загривок;
IV — встряхивание во время хватки за горло;
V — укладывание партнера на землю применением грубой силы;
VI — укладывание партнера на землю применением подсечки;
VII — схватывание пастью разных частей тела партнера;
VIII — завершение формирования элементов игры.
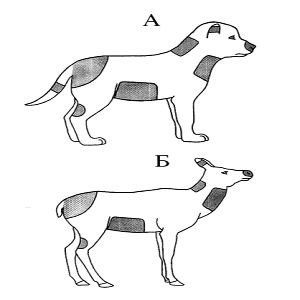
Во время игры в возрасте 40—50 дней формируется реакция хватания ртом различных частей тела партнера. Параллельно с этим необходимо привести материал, который выявляет основные атакуемые участки тела потенциальной жертвы как опытными хищниками, так и животными, не имеющими опыта охоты. Как видно из иллюстраций, атаки во время игры (рис. 14А) и во время охоты (рис. 14Б) направлены на одни и те же участки тела.