|
|
Популярные авторы:: Борхес Хорхе Луис :: Чехов Антон Павлович :: Толстой Лев Николаевич :: Грин Александр :: Горький Максим :: Азимов Айзек :: Раззаков Федор :: БСЭ :: Андреев Леонид Николаевич :: Joyce James Популярные книги:: The Boarding House :: Справочник по реестру Windows XP :: Детки в клетке :: Пауки :: Тайна похищения генерала Кутепова :: Дюна (Книги 1-3) :: Белолобый :: За волю (на белорусском языке) :: Тихие будни :: Я тебя никогда не забуду |
Волк. Вопросы онтогенеза поведения, проблемы и метод реинтродукцииModernLib.Net / Биология / Бадридзе Ясон / Волк. Вопросы онтогенеза поведения, проблемы и метод реинтродукции - Чтение (стр. 7)
В период с 3 по 10 мая 1977 года у реинтродуцированных нами животных появилось второе потомство. После их рождения группа снова воссоединилась, и старшие сибсы активно участвовали в выращивании волчат. Количество новорожденных детенышей удалось установить только после достижения ими возраста одного месяца. Оказалось, что в группе прибавилось 4 волчонка. Когда им исполнилось 3 месяца, мы снова провели анализ экскрементов со стандартным интервалом (n-73, n-85, n-70). Анализ показал, что все члены группы продолжали питаться только дикими животными. По остаткам пищи определили интервал между удачными охотами взрослых. Он составил 3—4 дня. В этом интервале все волки интенсивно охотились на грызунов.
В этот же период проверили прочность реакции избегания у волков относительно человека и возможность формирования этой реакции у новой генерации. Проведенные 60 экспериментов показали, что у всех реинтродуцированных нами животных реакция избегания сохранилась без всяких изменений. У старших сибсов реакция избегания относительно нас и незнакомых людей была четко дифференцирована. Нас они подпускали на расстояние 10—25 метров, а незнакомых людей — не менее, чем на недосягаемое для выстрела расстояние. У прибылых указанная реакция вырабатывалась за счет одноразового обучения, т. е. так же, как в свое время у их старших сибсов. И в этом случае хорошо была выражена дифференциация интенсивности реакции избегания относительно нас и незнакомых людей. В возрасте 7—8 месяцев погибли два щенка. Судя по внешним симптомам, причиной гибели была т. н. «собачья чума». Следует отметить, что, когда молодым животным исполнилось 8 месяцев, в группе возникли сильные агрессивные взаимодействия, в которых не принимали участия молодые особи. В результате возникшего конфликта из группы «выдворили» троих низкоранговых индивидов, из которых один — старший сибс (возраст 1 год 8 месяцев) и два — реинтродуцированные нами волка. В течение одного месяца не удавалось установить местонахождение этих волков, но затем их обнаружили между территориями I+ и II групп, в т. н. «буферной зоне». Надо отметить, что они часто пытались вернуться в свою группу, но оставшиеся в ней волки, включая молодых, встречали их сильной агрессивной реакцией. Изгнанные индивиды пытались также проникнуть на территорию соседней стаи, что также закончилось конфликтом. Именно во время конфликта с соседней стаей погибли два индивида, судьба третьего осталась неизвестной. II группа. После перехода четырех индивидов в первую группу и перераспределения территории данная группа заняла площадь в 65 км2. За этот период изменений в рационе группы не было. На третьей неделе беременности самка, а вместе с ней и самец отделились от стаи, но остались на той же территории. За неделю до родов мы обнаружили только одно логово. Установить точную дату родов волчицы не удалось из-за нашего отсутствия в этот период. Волчата родились в интервале между 15 апреля и 2 мая 1976 года. К этому периоду изменений в рационе волков не было. К моменту рождения волчат на территории стаи не было уже ни одной одичавшей собаки. Так же, как и в случае I группы, часть собак была убита волками, другая часть покинула данную территорию. Наблюдения за волчатами начали по достижении ими 3-месячного возраста. Оказалось, что к стае прибавилось 6 волчат. Трое погибли в возрасте 4 месяцев в результате заболевания «собачьей чумой». При первой попытке установления контакта с волчатами события развивались так же, как и в группе I+. В частности, самка издала описанный нами ранее лай тревоги, но сама осталась на месте. В ответ на этот лай у щенков появилась ярко выраженная реакция избегания. После этого случая, при каждой попытке контакта щенки активно избегали нас и наблюдали за нами с расстояния 15—30 метров. По-другому развивалась реакция волчат относительно незнакомых для родителей людей. Мы попытались спровоцировать встречу незнакомых людей со щенятами 38 раз. При первой же попытке у реинтродуцированных животных была реакция такой же интенсивности, как и перед реинтродукцией, т. е. фыркающий лай тревоги и убегание. Во всех последующих случаях волчата самостоятельно избегали людей. Дистанция избегания каждый раз была достаточной. Аналогично развивались события и при предъявлении домашних животных (30 случаев). И здесь, как и при формировании реакции избегания относительно человека, решающее значение имела реакция родителей. Анализ экскрементов, проведенный со стандартным интервалом (n-67, n-83, n-53) показал, что все волки питались только дикими видами. В период между 3 и 13 мая 1978 года у реинтродуцированных нами животных появилась вторая генерация. После рождения второй генерации группа заново воссоединилась 3, и старшие сибсы стали активно участвовать в воспитании детенышей. Определить число новорожденных удалось только после того, как им исполнился 1 месяц. Оказалось, что родились 4 волчонка. Когда им исполнилось 3 месяца, провели повторный анализ экскрементов со стандартным интервалом (n-88, n-78, n-73). Анализ показал, что все члены группы питались только дикими животными. В этот же период у волков проверили прочность реакции избегания человека и возможность ее формирования у нового поколения. Было проведено 35 экспериментов, которые показали, что у всех реинтродуцированных нами животных реакция избегания незнакомых людей была такой же, как и в момент реинтродукции. Родившиеся же уже на воле, избегали нас на расстоянии 10—25 метров, а незнакомых людей — на достаточно безопасное расстояние. У новой генерации эта реакция сформировалась в результате одноразового опосредованного обучения, т. е. так же, как и у их сибсов. Когда волчатам исполнилось 8 месяцев, в группе между старшими волками стали развиваться сильные агрессивные взаимодействия, в которых молодые не принимали участия. В результате возникшего конфликта из группы изгнали четырех реинтродуцированных нами низкоранговых индивидов. Последние поселились на смежной свободной территории, площадь которой через месяц составила примерно 40 км2. Следует отметить, что покинувшие группу индивиды в течение 2 недель пытались вернуться на прежнюю территорию, но каждый раз наталкивались на сильное агрессивное сопротивление. В процессе мечения «изгнанниками» границ новой, освоенной территории, сформировалась «буферная зона» вблизи с границами I и II групп. III группа. Реинтродукция осуществлена 10 января 1978 года вблизи деревни Зирта. Спустя месяц территория группы составили приблизительно 70 км2. Следует отметить, что в течение первой недели восточная граница территории этой группы непосредственно соприкасалась с границей II группы, из-за чего между членами двух групп постоянно отмечали сильные агрессивные взаимодействия. К концу первого месяца реинтродукции расстояние между границами групп составило от 1,5 до 3 км. С возникновением нейтральной зоны агрессивные взаимоотношения прекратились. В течение первого месяца в рационе группы изменений не было. В конце второй недели беременности доминантная самка и доминантный самец отделились от остальной группы и начали жить отдельно, оставаясь, однако, на той же территории. Уже за 2 недели до родов волки выбрали два пригодных для логова места. Волчата родились между 27 и 28 апреля 1978 года. Через 5 дней после их рождения группа снова воссоединилась, и ее члены стали охотиться вместе. После рождения первой генерации рацион волков остался без изменений. Так же как и в описанных выше случаях, к моменту рождения волчат на территории стаи уже не было ни одной одичавшей собаки — часть уничтожили волки, часть ушла за пределы данной территории. Наблюдать за волчатами мы начали по достижении ими 3-месячного возраста. Оказалось, что в стае прибавилось 6 волчат. Следует отметить, что при первой попытке контакта с детенышами, как и в случае с предыдущими группами, волчица издала фыркающий лай тревоги, но осталась на месте. В ответ на эти звуки у волчат появилась ярко выраженная реакция избегания. После этого случая щенки активно избегали нас и интенсивно наблюдали за нами с расстояния около 20—30 метров. Иначе развивалась реакция волчат на незнакомых для родителей людей. 42 раза мы пытались спровоцировать встречу щенят с незнакомыми людьми. При первой же попытке у реинтродуцированных нами животных проявлялась такая же реакция, как и перед реинтродукцией, т. е. фыркающий лай тревоги и убегание. Поведение родителей вслед за ними повторили и детеныши. Во всех последующих случаях волчата самостоятельно избегали незнакомых людей. Дистанция избегания всегда была достаточной. Логически схоже развивались события и при предъявлении волкам домашних животных (35 случаев). И здесь, как и при формировании реакции избегания человека у волчат решающее значение имела реакция родителей и подражание ей… После достижения волчатами 3-месячного возраста был проведен анализ экскрементов со стандартным интервалом (n-69, n-68, n-50). Анализ показал, что они питались только дикими животными. В период с 3 по 10 мая 1979 года у реинтродуцированных нами животных этой группы также появилась вторая генерация. После родов группа воссоединилась, и старшие сибсы принимали активное участие в воспитании волчат. Количество новорожденных удалось установить только спустя месяц после их рождения. Оказалось, что родилось четверо волчат. После достижения ими 3-месячного возраста были проведены анализы экскрементов со стандартным интервалом (n-43, n-70, n-57). Анализ показал, что все волки, как и прежде, питались дикими животными. В этот же период проверили прочность реакции избегания и возможность ее формирования у нового поколения. Было проведено 60 экспериментов, которые показали, что у всех реинтродуцированных животных реакция избегания сохранилась без изменений. У старших сибсов реакции на нас и на незнакомых людей были четко дифференцированы, как и во всех вышеуказанных случаях. Они избегали нас, начиная с расстояния 10—25 метров, незнакомых людей — с более безопасного расстояния. У нового поколения эта реакция выработалась в результате опосредованного обучения — в данном случае, в основном, на основе подражания реакциям старших сибсов. И в этом случае хорошо выражалась дифференциация реакции избегания нас и незнакомых людей. В период между 7 и 8 месяцем погибли два щенка. Судя по внешним симптомам, причиной смерти, опять таки, была т. н. «собачья чума». Следует отметить, что, когда молодым волкам исполнилось 8 месяцев, в группе (как и в вышеописанных случаях) возникли ярко выраженные агрессивные взаимодействия, в которых не участвовали молодые особи. В результате конфликта из группы были изгнаны 3 особи, из которых один — старший сибс в возрасте около 20 месяцев, и два — реинтродуцированных нами индивида. Через некоторое время изгнанных волков обнаружили в т. н. «буферных зонах», находящихся между территориями II и III групп. За 1,5 месяца нахождения здесь эти животные систематически пытались вернуться в прежнюю группу, но ее члены, включая молодых, встречали их сильной агрессией. Кроме того, они пытались вторгнуться и на территорию соседней группы, что также завершалось конфликтом. В конце концов «изгнанники» заняли свободную территорию северо-восточнее III группы. В течение 3 недель между территориями образовалась нейтральная зона шириной около 1—2,5 км. IV группа. Реинтродукция осуществлена 25 января 1979 года вблизи урочища Нахширгора. Через месяц после реинтродукции, территория группы составляла приблизительно 65 км2. Северная граница территории этой группы непосредственно примыкала к границам групп I+ и II, вследствие чего между членами IV, I+ и II групп развивались сильные агрессивные взаимодействия. К концу первого месяца реинтродукции расстояние между границами этих двух групп составило от 1-го до 3-х км. С возникновением нейтральной зоны агрессивные взаимодействия прекратились. В течение первого месяца рацион группы оставался без изменений. В конце второй недели беременности доминантный самец и доминантная самка отделились от остальной группы и начали жить отдельно, но на той же территории. Уже за 2 недели до родов волки выбрали два места, пригодных для логова. Точную дату рождения волчат установить не удалось из-за нашего отсутствия в этот период. По-видимому, волчата родились между 25 апреля и 5 мая 1979 года. После их рождения группа заново воссоединилась. Рацион волков не изменился. К моменту рождения волчат на территории стаи, как и во всех остальных случаях, не было ни одной одичавшей собаки. Наблюдать за волчатами начали по достижении ими 3-месячного возраста. Оказалось, что в группе прибавилось 6 волчат. Следует отметить, что при первой попытке контакта с детенышами события развивались так же, как в вышеописанных случаях. Затем щенки активно избегали нас на дистанцию около 10—20 метров. Как и в остальных случаях, заметна была четкая дифференциация между реакциями на нас и на незнакомых родителям людей. 30 раз мы пытались спровоцировать встречу щенят с незнакомыми людьми. При первой же попытке у реинтродуцированных нами животных проявлялась такая же реакция, как и перед реинтродукцией. Поведение родителей вслед за ними повторили и детеныши. Во всех последующих случаях волчата самостоятельно избегали незнакомых людей. Дистанция избегания всегда была достаточной. Логически схоже развивались события и при предъявлении волкам домашних животных (47 случаев). И здесь, как и при формировании реакции избегания человека у волчат решающее значение имела реакция родителей и старших сибсов. После достижения волчатами 3-месячного возраста был проведен стандартный анализ экскрементов (n-35, n-62, n-54). Анализ показал, что в их рацион не входили домашние животные. Через год, в период с 15 по 25 апреля 1980 года у реинтродуцированных нами животных IV группы появилось второе поколение. Старшие сибсы активно включились в воспитание детенышей. Количество новорожденных удалось установить только спустя 1,5 месяца. Оказалось, что родилось 5 волчат. После того, как им исполнилось 3 месяца, провели серию стандартных анализов экскрементов (n-65, n-82, n-40), который показал, что и все эти волки также питались только дикими животными. В этот же период, как и всегда, проверили прочность реакции избегания человека и возможность ее формирования у нового поколения. Было проведено 30 экспериментов, которые показали, что у всех реинтродуцированных животных реакция избегания сохранилась без изменений. У старших сибсов реакция на нас и на незнакомых людей были четко дифференцированы, как и во всех вышеуказанных случаях. Они избегали нас на расстоянии 10—25 метров, незнакомых людей — на достаточное расстояние. У нового поколения эта реакция выработалась по такому же принципу, что и во всех предыдущих случаях — в результате одноразового опосредованного обучения. Как видно из представленного материала, скорость выработки реакции избегания, ее дифференцированность и прочность во всех группах были одинаковыми. В связи с тем, что во всех реинтродуцированных нами группах явления развивались одинаково, можно даже сказать, стереотипно, на данном этапе наблюдений мы сочли работу завершенной. Надо отметить, что кратковременные наблюдения за отделившимися группами и их потомством (около 50 часов на каждую группу) показали, что у всех животных, включая потомство, полностью сохранена реакция избегания людей и домашних животных. Анализ экскрементов, проведенный два раза в течение года (всего 115 анализов), показал отсутствие домашних животных в рационе волков. Проводимые ежегодно опросы (по 1987 год включительно) местного населения и пастухов не выявили фактов хищничества волка в отношении домашних животных. Отмечены были три случая хищничества медведя. Ни одного случая возврата ошейников реинтродуцированных нами волков не было. Полученный материал однозначно указывает на то, что все 22 реинтродуцированных нами волка в течение всего периода наблюдений активно избегали незнакомых людей и не нападали на домашних животных. Более того — они целенаправленно обучали свое потомство тем навыкам, которым в свое время мы обучили их. Необходимо подчеркнуть и то, что старшие сибсы так же целенаправленно обучают младших тем навыкам, которым они сами обучились у родителей. Таким образом, все 35 волков выживших из 40 прибылых, активно избегали незнакомых людей и не нападали на домашних животных. Надо специально отметить, что одним из решающих условий успеха реинтродукции волков было участие представителей местного населения в этом процессе, что полностью сняло проблему их антагонизма к реинтродуцируемому виду. Это произошло потому, что реакцию избегания людей и домашних животных мы вырабатывали при их непосредственном участии в результате чего люди убеждались в том, что волк может стать неопасным для них самих и их собственности. Для успешной охоты, помимо формирования ее элементов в процессе постнатального онтогенеза, животным необходимо детальное знание территории, т. е. формирование т. н. «мысленной карты», которая обеспечивает свободное ориентирование на ней — животному необходимо знать, как и где обнаружить добычу, затаиться или просто перемещаться в нужном направлении. Разработанный нами метод дает возможность решить и эту задачу. С учетом полученного нами материала и специфики высшей нервной деятельности, этот метод может быть применен к любому виду крупных наземных млекопитающих. Таким образом, становится возможным решение проблемы восстановления исчезающих или исчезнувших в природе видов путем реинтродукции в естественную среду обитания выращенных в неволе индивидов.
Глава 3.2. Реакция диких копытных на хищных: — инстинкт или традиция?
|
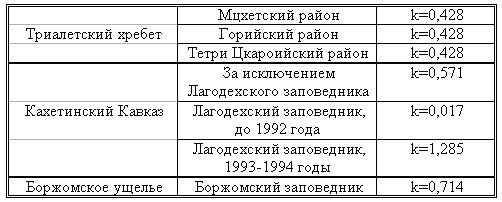
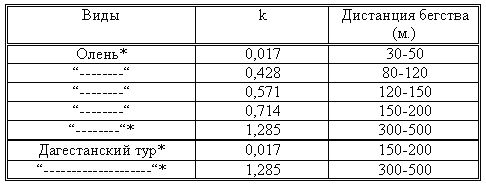
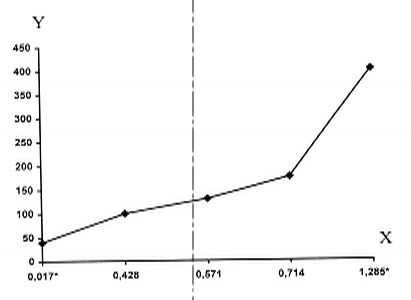
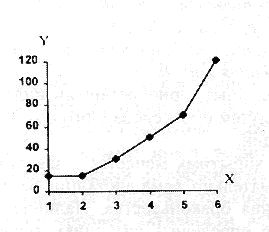
|||||||