З.А. Зорина, Л.В. Крушинский, Л.Г. Романова, И.И. Полетаева
Основные положения концепции Лоренца
Таким образом, факты, первоначально использованные Лоренцем, были в той или иной мере известны и более ранним исследователям. Лоренц, создавая свою «рабочую гипотезу поведения», опирался не только на собственный экспериментальный материал, но творчески осмыслил и привел в единую систему представления своих предшественников. Большое значение имел также тот факт, что Лоренц уточнил применявшуюся для описания поведения терминологию. Он ввел ряд новых понятий и уточнил некоторые из принятых его предшественниками терминов.
В качестве единицы инстинктивного поведения Лоренц выделяет «наследственные координации», или, как он позднее называл их «эндогенные движения», т. е. видоспецифические (одинаковые у всех особей данного вида), врожденные (т. е. проявляющиеся в «готовом виде», без предварительной тренировки) шаблонные (т. е. стереотипные по порядку и форме исполнения) двигательные акты. Сейчас их принято называть комплексами фиксированных действий (Fixed action patterns).
В основу своей первоначальной концепции Лоренц положил деление поведения на врожденное (собственно инстинктивное) и приобретенное (сформированное за счет индивидуального опыта, обучения). Он указывал, что такое дробление в большинстве случаев бывает условным. Каждая последовательность поведенческих актов рассматривается Лоренцом как сцепление инстинктов и обучения. Подробнее вопрос о проблеме врожденного и приобретенного в трудах этологов рассматривается в гл.5. Наследование видоспецифических особенностей в выполнении фиксированных Комплексов действий можно анализировать, изучая поведение гибридов первого поколения от скрещивания особей родственных видов, у которых это поведение четко различается, а также (что применимо в основном к насекомым) у особей с локальными мутациями, затрагивающими этот признак (см. гл.7). Первыми в этом отношении были эксперименты Уитмена и Хейнрота по скрещиванию близких видов горлиц и уток, обнаружившие специфику наследственной обусловленности фиксированных комплексов действий при ухаживании и демонстративном поведении. Для выяснения роли индивидуального опыта в их проявлении применялись эксперименты с воспитанием молодых животных в изоляции от действия определенных факторов внешней среды.
Структура поведенческого акта
Рассмотрим, каким образом Лоренц представлял себе структуру целостного поведенческого акта.
Поисковое поведение.
Как уже указывалось, поведение животного — это не всегда пассивная реакция на внешние раздражители. Во многих случаях животное, пришедшее в состояние специфической готовности к какому-то виду деятельности (например, готовности к размножению), активно ищет стимулы, при действии которых эта деятельность могла бы осуществиться. Так, в начале сезона размножения самцы территориальных видов птиц выбирают место для гнезда и охраняют занятый участок, ожидая появления самки. У ряда видов, образующих пары лишь на один сезон, самец в начале весны должен разыскивать самку. Вслед за Крэгом Лоренц называет эту фазу поведенческого акта поисковой (или аппетентной — appetitive). Поисковое поведение представляет собой вариабельный комплекс реакций и характеризуется «спонтанностью», (так как оно проявляется главным образом под влиянием внутренних стимулов) и вариабельностью выполняемых во время нее движений.
Окончание этой фазы наступает, когда животное достигнет ситуации, в которой может осуществиться следующее звено данной цепи реакций.
Например, выбор гнездовой территории иногда ограничивается перелетом в определенное, ранее уже использованное место, а в других случаях требует и длительных поисков, борьбы с другими самцами, а при поражении — выбора нового участка. По представлению Лоренца, именно поисковая фаза поведенческого акта относится к категории целенаправленного поведения, так как совершаемые действия подчинены определенной цели, которая может быть достигнута разными путями.
Поисковая фаза, как и завершающий акт, строится на врожденной основе, но в ходе онтогенеза эта основа дополняется приобретенными реакциями. Именно поисковое поведение является средством индивидуального приспособления животных к окружающей среде, причем это приспособление бесконечно разнообразно по своим формам. Основу формирования поискового поведения в онтогенезе составляют такие процессы, как привыкание и обучение во всех его многообразных формах. Именно к поисковой фазе поведенческого акта относятся и проявления элементарной рассудочной деятельности животных, когда для достижения цели животное в новой для него ситуации оперирует ранее сформировавшимися у него понятиями и уловленными им эмпирическими законами, связывающими предметы и явления внешнего мира (Крушинский, 1977). В рассматриваемом нами примере роль обучения может проявиться, например, в том, что участки и места для гнезд, выбираемые взрослыми опытными самцами, как правило, лучше, чем у птиц, впервые участвующих в размножении.
Простая схема «поисковое поведение — ключевые стимулы — завершающий акт», которую обычно используют для описания поведения, оказывается включенной в более сложную систему. Так, например, в некоторых случаях поисковое поведение приводит не к завершающему акту, а к такому сочетанию раздражителей, которое стимулирует следующую фазу поискового поведения. Например, весной с наступлением тепла у колюшки развивается поисковое поведение — выбор территории для гнезда. Когда найдена подходящая территория, начинается следующая стадия поискового поведения — строительство гнезда, а затем ухаживание за самкой. Для определения таких видов многостадийного поискового поведения ученик Тинбергена Берендс ввел понятие «иерархии поискового поведения».
Завершающий акт.
В отличие от вариабельного по форме поискового поведения непосредственное осуществление стоящей перед животным цели, удовлетворение руководившего им побуждения происходит в виде видоспецифических фиксированных комплексов действий. Они лишены приобретенных элементов и могут совершенствоваться в онтогенезе только за счет созревания ответственных за них структур мозга, но не за счет обучения. Типичные примеры таких действий — различные формы угрожающего и полового поведения. Подчеркивая тот факт, что реакции типа завершающих актов у всех представителей вида (или более крупной систематической категории) одинаковы и проявляются у молодых животных без специального обучения, Лоренц назвал их эндогенными движениями. Именно, реакции типа завершающих актов и представляют собой, по Лоренцу, инстинкт в чистом виде, как он был определен выше. Как уже указывалось, такие реакции могут быть более устойчивыми в филогенезе, чем многие морфологические признаки. Примером этого служит одновременное вытягивание крыла и ноги, а также шеи и крыла, наблюдаемое у всех птиц.
Значение понятия о завершающем акте для изучения эволюции поведения.
Введение понятия о завершающем акте было плодотворным прежде всего для изучения эволюции поведения. Оно позволило применить к изучению поведения принцип гомологизации, заимствованный из сравнительной анатомии, и успешно использовать его в систематике. Рассматривая стабильные видоспецифические формы инстинктивного поведения, Лоренц, вслед за Уитменом, указывал, что зачастую они оказываются более древними и охватывают более широкую группу животных, чем некоторые морфологические признаки, на которых основывается систематика, Например, у ночной цапли рода Nycticorax церемония приветствия аналогична таковой у южноамериканской цапли рода Cochlearious, что указывает на их общее происхождение. Морфологическая же структура, связанная с осуществлением этой церемонии, — оперение головы — устроена у этих видов совершенно по-разному, что свидетельствует о ее более позднем возникновении. В одной из своих статей Лоренц показал, насколько расширяются возможности систематики, если в качестве таксономических признаков использовать особенности поведения. Они позволят, в частности, дифференцировать виды, совершенно не различающиеся морфологически. Вскоре Лоренц подтвердил этот тезис, изучив церемонии ухаживания у 16 видов уток (см. Хайнд, 1975). Оказалось, что некоторые признаки, например «односложный поиск» у птенца и «реакция питья как сигнал мир», характерны для всех видов отряда утиных. Некоторые виды и семейства отличаются друг от друга как по морфологическим, так и по поведенческим признакам. Например, гусеобразные отличаются от прочих утиных как по характеру писка подросшего птенца, так и по однотонности их окраски. Между тем в других случаях поведенческие признаки служат единственным достоверным отличием. Например, у мускусных уток перед спариванием наблюдаются характерные движения головы, как будто бы птица «нацеливается» клюнуть другую. Такая форма поведения не наблюдается у родственных мускусной утке видов.
Вслед за Лоренцом другие исследователи подобно проанализировали многие систематические группы. В качестве примера можно указать исследование поз угрозы и ухаживания у 15 видов пауков Sallicidae и оборонительных поз у 15 видов богомолов, исследование поведения цихлидовых рыб, и целый ряд других исследований как на позвоночных, так и на беспозвоночных животных. Этологический анализ поведения большого числа видов семейства кошачьих — в том числе формирования охотничьих навыков и взаимодействий животных друг с другом в сопоставлении с данными по анатомии и экологии — позволил П. Лейхаузену (Leyhausen, I979) пересмотреть классификацию этого семейства и внести в нее некоторые изменения. Крупный вклад в изучение эволюционных аспектов поведения внес Тинберген и его ученики.
Врожденный разрешающий механизм.
Второе важное положение концепции Лоренца опиралось на тот факт, что многие инстинктивные действия проявляются только в ответ на определенные раздражители, которые были названы ключевыми или знаковыми. Характерно, что эти раздражители животные опознают уже при первом предъявлении, без всякого индивидуального опыта. Например, красное пятно на брюшке рыбы обязательно вызывает агрессивную реакцию у любого самца трехиглой колюшки, так как он начинает воспринимать такую рыбу как соперника. Для самца определенного вида птиц в начале весны ключевыми будут стимулы, характерные для самки их вида. Эти стимулы снимают блокирующие механизмы в нервной системе самца и тем самым способствуют проявлению соответствующей инстинктивной реакции. Этот механизм снятия блока Лоренц назвал «врожденной схемой реагирования» (angeborne auslosende Schema) в настоящее время более распространен предложенный английскими этологами термин «врожденный разрешающий механизм» (innate releasing mechanism). Для пояснения принципа его действия часто пользуются аналогией с ключом и замком. Знаковый стимул сравнивают с ключом, идеально соответствующим замку — врожденному разрешающему механизму, который снимает торможение с центров, обеспечивающих осуществление соответствующей реакции.
Ключевые раздражители.
Раздражители, при действии которых происходит срабатывание «врожденного разрешающего механизма», получили название ключевых, или знаковых. Они весьма разнообразны по своей природе и могут быть адресованы любому из анализаторов. Специфические вещества — половые аттрактанты, феромоны, улавливаемые готовыми к размножению половыми партнерами, выступают как ключевые раздражители в половом поведении многих насекомых, амфибий и ряда млекопитающих. У многих животных и птиц в качестве ключевых стимулов полового поведения выступают морфологические признаки — особенности окраски тела, специальные структуры (например, гребни и хохолки у птиц). «Зеркальца» на маховых перьях селезней многих видов уток также являются одним из ключевых раздражителей в половых демонстрациях. Этот вид зрительно воспринимаемых ключевых стимулов принято называть «релизерами». Видоспецифические звуковые стимулы — пение, крики угрозы или ухаживания — также будут ключевыми раздражителями для фиксированных комплексов инстинктивных действий у животных разных видов. Особую категорию ключевых стимулов составляют видоспецифические комплексы движений. Примером могут служить брачные демонстрации, позы угрозы и подчинения, приветственные ритуалы и т. д. Подробное изложение современных представлений о врожденном разрешающем механизме можно найти в книге Меннинга (1982).
Запечатление.
Особую группу ключевых стимулов составляют такие, для опознавания которых требуется специфический тип обучения — запечатление. В отличие от обычных форм обучения, происходящего на протяжении всей жизни животного, эта форма приобретения индивидуального опыта приурочена к строго определенному «чувствительному» периоду онтогенеза. В этот период животное «фиксирует», «запечатлевает», раздражитель, на который впоследствии будет осуществляться та или иная врожденная реакция. Это явление было известно биологам, но Лоренц первый полно описал, а главное — определил его роль в формировании поведенческого акта.
Классический пример запечатления — формирование реакции следования за матерью у птенцов выводковых птиц, или, по современной терминологии, запечатление привязанности. Сама по себе реакция эта врожденная, но в течение первых часов после вылупления молодые птицы должны «запечатлеть» облик матери. Если в этот период утята не увидят, утки, то впоследствии будут ее бояться. Более того, у утенка, не встретившего в соответствующий период подходящего для запечатления объекта, в дальнейшем его поведение будет резко нарушено: он будет избегать контактов со всеми живыми существами. Если же в этот период на глаза утенку попадает какой-нибудь движущийся (даже неодушевленный) предмет, то он начинает реагировать на него как на свою мать и всюду следовать за ним. Так, например, Лоренц выращивал разных представителей отряда гусеобразных в изоляции от сородичей в течение первой недели жизни и отметил, что такие птицы предпочитают следовать за людьми, а не за птицами своего вида. Многие из них постоянно держались перед дверью дома Лоренца и пытались следовать за людьми, как только те выходили из дома.
Лоренц указывал, что по своим свойствам запечатление существенно отличается от обычного ассоциативного обучения прежде всего тем, что оно происходит в определенный, достаточно узко ограниченный период онтогенеза. Второе его отличие состоит в том, что эффект запечатления необратим и в обычных условиях не угашается. В связи с этим запечатление птицей особи чужого вида может полностью дезорганизовать ее общественное поведение в будущем. Характерно также, что реакция следования обнаруживается не только при виде конкретной особи (или предмета), которых запечатлела в соответствующий период молодая птица, но и любых особей того же вида или предметов того же типа, что и объект запечатления.
Анализируя явление запечатления и его роль в формировании поведения, Лоренц обращал внимание на сходство его с процессом дифференцировки органов в эмбриогенезе. В обоих случаях наличие чувствительного периода — необходимое условие для осуществления определенной реакции формообразования. Наиболее подробно эта проблема изложена в монографии Лоренца «Эволюция и модификации поведения» (1965) (Evolution and Modification of Behavior).
Помимо запечатления привязанности, которое обеспечивает контакт птенцов с матерью в ранний период онтогенеза, существует и так называемое половое запечатление, которое обеспечивает последующий правильный выбор полового партнера птицей по достижении ею половозрелости. Если первая форма запечатления характерна для выводковых птиц, то вторая форма играет существенную роль и в формировании поведения птенцовых. Так, например, в опытах Лоренца и его учеников селезни, воспитанные уткой другого вида, став взрослыми, пытались спариваться только с самками вида-воспитателя, игнорируя самок собственного вида. Сходным образом ведут себя и птенцовые виды. Самцы зебровой амадины, воспитанные бенгальскими амадинами, впоследствии ухаживали только за самками вида-воспитателя.
Описав явление запечатления, Лоренц привлек к нему пристальное внимание исследователей многих специальностей, которые изучили его роль в формировании поведения животных разных систематических групп, уточняли продолжительность и значение чувствительных периодов, влияние параметров запечатлеваемого стимула, сопоставляли свойства запечатления и ассоциативного обучения и т. д. Подробный разбор современного состояния этой проблемы можно найти в книге О. Меннинга (1982).
Спонтанность.
Третьим компонентом концепции Лоренца является тот факт, что наряду со строго рефлекторным проявлением многих инстинктивных действий в ответ на ключевой стимул в ряде случаев они проявляются спонтанно. Под спонтанностью принято понимать такие изменения на выходе системы, для которых неизвестны соответствующие изменения на входе (Хайнд, 1975).
Классический пример спонтанного проявления инстинктивного действия, которое в обычных условиях возникает в ответ на специфический стимул, — наблюдение Лоренца за поведением ручного скворца, который был воспитан в изолированных условиях и не имел возможности ловить на лету насекомых, т. е. выполнять стереотипную видоспе-цифическую реакцию — заключительный акт пищедобывательного поведения. Оказалось, что птица стала все более неразборчиво реагировать на внешние раздражители, пытаясь преследовать их, что свидетельствует о снижении порога реакции. Если адекватные раздражители — насекомые — так и не появлялись, то в их отсутствие наблюдалась полная последовательность движений, необходимых для их поимки. Скворец неожиданно взлетал, щелкал клювом в пустоте, возвращался на жердочку, выполнял характерные движения, которыми обычно умерщвляется насекомое, и, наконец, «глотал муху». По выражению Лоренца, реакция происходила в этом случае «вхолостую». Подобно этому Хайнд впоследствии описал, как канарейка, лишенная материала для строительства гнезда, переносила и вплетала несуществующие травинки в несуществующее гнездо. Эти примеры иллюстрируют возможность существенного снижения порога инстинктивных действий, если они долго не совершались в силу каких-то причин.
Наряду с этим для многих случаев характерно повышение порога реагирования, когда соответствующая реакция воспроизводится с трудом или вообще не воспроизводится. Например, в начале сезона размножения брачные позы и демонстрации самцов вызывают ответную реакцию только у самок, уже готовых к размножению, тогда как, например, У молодых самок они не вызывают никаких реакций. Вместе с тем зимой брачные демонстрации перестанут быть эффективными даже для взрослых самок.
Последующий анализ показал, что все сложные поведенческие акты в той или иной степени содержат как спонтанные, так и рефлекторные элементы. В действительности очень трудно бывает установить, до какой степени отдельные элементы поведения спонтанны или же вызываются внешними стимулами, которых просто не может уловить наблюдатель. Далее, постоянные внешние условия сами по себе через некоторое время могут становиться внешними стимулами для возникновения соответствующих реакций. Вопросы спонтанности и ее роли в поведении впоследствии были подробно исследованы рядом авторов. Изложение этой проблемы можно найти в монографии Хайнда (1975) и в книге Меннинга (1982). Мы укажем лишь, что этот факт сыграл важную роль в формировании представлений Лоренца о внутренних механизмах поведенческого акта.
Гипотеза Лоренца о внутренних механизмах инстинктивных действий
На основе подобных фактов о свойствах инстинктивных действий Лоренц выдвинул ряд положений об их внутренних механизмах. Согласно его представлениям, под действием ряда внешних и внутренних факторов (гормоны, температура, освещенность и т. п.) в соответствующих нервных центрах происходит накопление «энергии или потенциала действия», специфических в отношении определенного побуждения (голод, жажда, половая потребность и т. п.). Возрастание этой активности выше некоторого уровня приводит к проявлению поисковой фазы поведенческого акта, которая, как уже говорилось, характеризуется широкой изменчивостью исполнения как у данной особи, так и у разных представителей одного вида. Она состоит в активном поиске раздражителей, при действии которых может быть удовлетворено возникшее у животного побуждение. Когда эти раздражители найдены, осуществляется завершающий акт — фиксированный комплекс видоспецифических движений, стереотипных как у одной особи в каждом случае, так и у всех особей данного вида. Этот комплекс движений характеризуется высокой степенью генотипической обусловленности. При усиленном накоплении «специфической энергии действия» завершающий акт может осуществиться спонтанно, т. е. в отсутствие соответствующих раздражителей (реакция «вхолостую»). Термин «специфическая энергия действия» применялся в значительной мере как метафора и должен был подчеркнуть, что внутренние мотивирующие факторы влияют только на определенные системы поведенческих реакций, связанные, например, с добыванием пищи и не связанных с размножением.
Нейрофизиологическая база представлений К. Лоренца
Следующий шаг, который сделал Лоренц и развили его последователи, — это попытка соотнесения гипотетической «специфической энергии действия» с имевшимися в тот период данными о реально происходящих в нервной системе процессах. Большую роль сыграли при этом данные о наличии автоматических спонтанных процессов в центральной нервной системе (цнс).
В тот же период, когда Лоренц (1937), исходя из наблюдений за поведением животных, постулировал представление о внутренних факторах, определяющих характер их проявления, немецкий физиолог Э. фон Хольст собрал и обобщил факты, которые стали нейрофизиологической базой концепции Лоренца.
Первоначально этологи пытались использовать для объяснения получаемых данных рефлекторную теорию, однако постепенно накапливалось все больше фактов, которые не могли быть объяснены с ее помощью. В соответствии с классической рефлекторной теорией всякое поведение является реакцией на внешний или внутренний стимул. Афферентный нерв проводит возникающее в рецепторе возбуждение в спинной мозг, в сером веществе которого находится соответствующий «рефлекторный центр». Далее возбуждение передается на эфферентный нерв. Эти три элемента составляют так называемую «рефлекторную дугу». Более сложные цепи поведения — это цепи отдельных рефлексов. Согласно этой теории, например, у угря при плавании сокращение и расслабление одного мышечного сегмента туловища механически возбуждает проприрецепторы следующего сегмента, а они в свою очередь возбуждают его рефлекторную дугу, что вызывает сокращение мышц этого сегмента. Однако фон Хольст, изучавший плавание угря обнаружил, что эта гипотеза не выдержала экспериментальной проверки. До фон Хольста наиболее распространенным являлся следующий экспериментальный метод: искусственно деафферентировали цнс, а затем путем периферической стимуляции вызывали периферическую реакцию, которая и доказывала, что цнс функционирует как рефлекторный механизм.
Фон Хольст изменил эту методику и сосредоточил внимание на активности самой цнс. Он показал, что в изолированной брюшной нервной цепочке земляного червя наблюдаются разряды ритмических импульсов, точно соответствующие сокращению сегментов ползущего червя. В дальнейшем, исследуя механизм плавания у угря, фон Хольст прибег к этому же методу. Если фиксировать средние сегменты тела угря, не давал им сокращаться, то, согласно рефлекторной теории, задние сегменты не должны придти в движение, не получив раздражения от соседних сегментов. Однако на самом деле через промежуток времени, соответствующий несостоявшемуся сокращению фиксированных экспериментальных сегментов, задние сегменты тела также приходят в движение. Это может быть показано еще более четко, если перерезать дорсальные корешки спинного мозга, передающие сенсорные импульсы. Оказалось, что несмотря на деафферентацию угорь сохраняет способность к хорошо координированным плавательным движениям. Это должно означать, что все плавательные движения полностью запрограммированы в цнс и выполняются независимо от внешних раздражений, а не по механизму рефлекторной дуги.
Существовал и ряд данных, которые лишь с натяжкой укладывались в рамки рефлекторных представлений. Было обнаружено, например, что у децеребрированных кошек две полностью деафферентированные антагонистические ножные мышцы ритмически сокращаются. Это привело к выводу, что в цнс существует эндогенный автоматизм, не зависящий от внешних раздражений. Позднее Вейсс пересадил участок спинного мозга и зачаток конечности у зародыша аксолотля. Развивающийся зачаток иннервировался пересаженным эмбриональным спинным мозгом. Поскольку мотонейроны развиваются быстрее сенсорных нейронов, их отростки, прорастая, достигают пересаженной конечности раньше, чем отростки сенсорных клеток. В тот момент, когда произошло афферентное соединение, конечность начала двигаться. Хотя совершаемые ею движения и не были полностью характерными для ходьбы, тем не менее в них можно было обнаружить примитивные формы координации, включая переменное сокращение мышц антагонистов. Этот факт также демонстрирует наличие эндогенного автоматизма в цнс и существование морфологических структур, обеспечивающих его проявление.
Аналогичные явления были продемонстрированы и на других позвоночных. Деафферентированные головастики, жабы и рыбы сохраняли способность плавать, нормально координируя движения, до тех пор, пока у них оставался неразрушенным хотя бы один чувствительный нерв. Следовательно, для поддержания плавательных движений необходим минимальный уровень афферентной импульсации, которая способствует поддержанию уровня возбуждения в соответствующих нервных центрах (или «специфической энергии действия» по Лоренцу).
Большое значение в этом плане имели работы Рёдера, который и обнаружил недостаточность рефлекторной теории для объяснения некоторых сторон поведения насекомых.
В последующие десятилетия эти данные о возможности спонтанной активности в цнс насекомых получили широкое экспериментальное подтверждение. По представлениям Рёдера, нервные элементы, лежащие в основе спонтанного и рефлекторного поведения, различаются, в частности, по порогу возбудимости. Порог раздражения первых может быть настолько низким, что инстинктивная реакция способна осуществляться помимо видимых внешних раздражителей.
Влияние уровня возбуждения в соответствующих нервных центрах на характер протекания инстинктивных реакций было подтверждено работами немецких ученых Э. фон Хольста и У. Сент-Поль (1960). В зависимости от локализации раздражаемой током структуры ствола мозга кур они наблюдали либо элементарные поведенческие реакции (клевание, повороты головы), либо сложные акты поведения (ухаживание за отсутствующими самками). Сложные реакции протекали менее константно, чем простые. Они зависели от состояния животного, от предшествующих стимулов от параметров тока и всей совокупности действующих на птицу стимулов. Например, при слабом токе петух бросался на чучело хорька, на которое до этого не обращал внимания, а при усилении тока такая же реакция проявлялась на неспецифический раздражитель — кулак.
Гидравлическая модель.
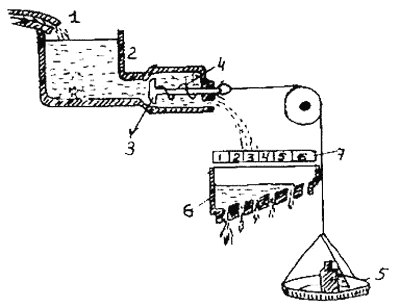
Лоренц предложил гипотетическую модель осуществления реакций типа завершающих актов, общие принципы которой он заимствовал из гидравлики. В настоящее время гидравлическую модель можно считать глубоким анахронизмом в этологии, представляющим лишь исторический интерес. Ввиду этого мы остановимся на ней очень кратко (рис.1).
Накопление «специфической энергии действия» представлено в модели в виде постепенного накопления воды в резервуаре.
Рис. 1. Схема гидравлической модели для объяснения регуляции поведения (см. Меннинг, 1982)
1 — кран, 2 — резервуар, 3 — клапан, 4 — пружина, 5 — чашка весов, 6 — лоток 7 — шкала
Давление жидкости внутри него все время повышается. Отток жидкости из резервуара (аналогизируемый с активностью животного) возможен по трубам, которые блокируются клапаном. Этот клапан может открыться при достаточно большом давлении (накоплении жидкости в резервуаре), а также при воздействии извне — при действии тяжести гири, приложенной к клапану. Такое внешнее воздействие аналогизируется с влиянием внешних раздражителей, специфичных для данного поведенческого акта. Нарастающее давление и действие внешней силы направлены одинаково и могут суммироваться. Чем больше жидкости набралось в резервуаре, тем меньше усилий нужно приложить извне, чтобы открыть клапан, а иногда клапан может открыться и без внешнего воздействия. Этот момент моделирует появление активности «вхолостую», возникающей без видимых внешних сигналов. Эта аналогия может быть продолжена. Если клапан открылся лишь немного, жидкость будет вытекать слабо и попадет только в область нижнего отверстия специального наклонного лотка. Если же клапан откроется сильнее — жидкость потечет сильнее и попадет и на следующие отверстия. Этот лоток с отверстиями аналогизируется с разными типами двигательной активности, относящейся к данному поведенческому акту. Самое нижнее отверстие соответствует активности с наиболее низким порогом — например, некоторым проявлением поискового поведения. Другие отверстия соответствуют иным формам активности, обладающим более высоким порогом. Если резервуар опорожняется, т. е. иссякает; «специфическая энергия действия», — данное поведение больше продолжаться не может.
Современная оценка гидравлической модели Лоренца.
В определенных пределах предложенная Лоренцем модель хорошо описывает феноменологию инстинктивных действий, а также циклические изменения, происходящие в реактивности нервной системы животного к внешним раздражителям — снижение порога совершения действия, если оно долго не выполнялось, восстановление готовности к инстинктивным действиям после перерыва и возможность появления реакций на неспецифические раздражители. В течение некоторого времени она являлась стимулом к развитию новых исследований. Однако наряду с этим модель Лоренца неоднократно подвергалась критике, отчасти необоснованной, но отчасти и справедливой (например, механистичность построения). При создании модели принимаются в расчет лишь самые общие черты, отражающие схему работы моделируемой системы, а также ее соответствие известным из опыта фактам. Другое дело, что предложенная Лоренцем модель, хорошо объясняя очень многие из известных фактов, объяснить другие была не в состоянии. Однако это свидетельствовало лишь о том, что представление о запасании «специфической энергии действия» имеет ограниченное применение, и соответствующие термины и понятия не универсальны.
Анализ применимости модели Лоренца к описанию поведения в сопоставлении с другой моделью, предложенной Дейчем, можно найти в книге Меннинга (1982). Плодотворность применения модели Лоренца мы иллюстрируем в гл.5 на примере анализа поведения кошек, выполненного Лейхаузеном.
Современная оценка концепции Лоренца о врожденном разрешающем механизме опирается на большое количество экспериментальных фактов, полученных в последние годы. Подробный обзор современных взглядов по этим вопросам приводится в книге Меннинга.
Очевидно, что и «специфическая энергия действия» и ключевые стимулы — это понятия, которые в переводе на язык современной нейрофизиологии выражаются такими терминами, как «специфическое побуждение», активация той или иной мотивационной системы, а также видоспецифическая избирательность перцепторного аппарата.
Развитие концепции К. Лоренца в работах Тинбергена
Представления Лоренца, заложившего основы этологии, развил голландский ученый Н. Тиберген. В 50-е годы Тинберген был приглашен в Оксфордский университет, где и была проведена большая часть его исследований. Работы Тинбергена и его учеников образовали особое направление, которое известно как английская школы этологии.
Тинбергену принадлежит разработка иерархической модели поведения, которая в большей степени учитывала физиологические данные, чем исходная модель Лоренца. На базе этой модели он выделил некоторые формы конфликтного поведения и высказал гипотезу о их механизмах.
Тинберген и его ученики в течение многих лет систематически исследовали в природных условиях поведение ряда видов насекомых и птиц. Классическим объектом их лабораторных исследований стала трехиглая колюшка — легко содержащийся в неволе вид пресноводных рыб. Репродуктивное поведение колюшки послужило моделью для выявления многих важных принципов организации поведения животных. Особое значение приобрели их работы по колониальным морским птицам. Они явились основой многих современных представлений о сообществах животных и факторах, регулирующих их структуру, а также о коммуникативном значении ряда поведенческих актов. Кроме того, они способствовали изучению проблемы многообразных форм приспособления животных к борьбе с хищниками, которая накладывает отпечаток практически на все стороны поведения. Впоследствии эта проблема изучалась и на других животных, Последовательное применение сравнительного метода и остроумные полевые эксперименты способствовали выделению приспособительного значения поведенческих признаков и их эволюционного происхождения. Рассмотрим основные направления работ Тинбергена.
Иерархическая теория инстинкта
Основой для разработанной Тинбергеном модели поведения послужили следующие факты. Известно, что между различными стереотипными двигательными реакциями существует ряд закономерных отношений. В некоторых ситуациях группы инстинктивных движений появляются совместно — они характеризуют определенное внутреннее состояние животного и проявляют общие флюктуации порога поведенческой реакции. Повышение порога реакции А поднимает порог реакции В (и наоборот), а это свидетельствует о том, что обе они зависят от общего функционального «центра». Наблюдая за сложными поведенческими комплексами действий, можно видеть некоторую регулярность в последовательности проявления тех или иных действий. В качестве примера можно привести агрессивные столкновения рыб за раздел территории. У многих костистых рыб, в том числе и у цихлид, им почти всегда предшествует демонстрация запугивания. Причем у одних видов эти столкновения следуют за очень коротким периодом запугивания; а у других — за весьма разнообразными демонстрациями запугивания, серьезное агрессивное столкновение с ранениями следует лишь в том случае, если силы обоих самцов равны. Наконец, у третьей группы видов настоящие драки уже не наблюдается, и крайне ритуализированная церемония запугивания выполняется до полного истощения одного из соперников, что и решает спор.
В такого рода ритуализованных столкновениях имеется специфическая последовательность движений: они начинаются с демонстрации боковых поверхностей тела, за которой следует подъем вертикальных плавников. Затем следуют удары хвостом, которые через посредство боковой линии, воспринимающей изменение давления воды, могут, вероятно, сообщить о силе противника. После этого противники встают друг перед другом, вслед за чем начинаются взаимные толчки с открытой пастью, а у других видов — укусы в открытый рот. Они продолжаются до тех пор, пока один из соперников не устанет, окраска его бледнеет и в конце концов он уплывает.
Такие ритуализованные драки и агрессивные столкновения — прекрасные примеры специфической последовательности стереотипных двигательных реакций: удары хвостом не начнутся до подъема спинного плавника, а толчки отмечаются только после многих ударов хвостом. По интенсивности демонстрации запугивания и ударов хвоста, опытный наблюдатель может определить, кто победит, и начнутся ли толчки «с открытой пастью» вообще, или же один из соперников просто сбежит до начала «серьезной драки».
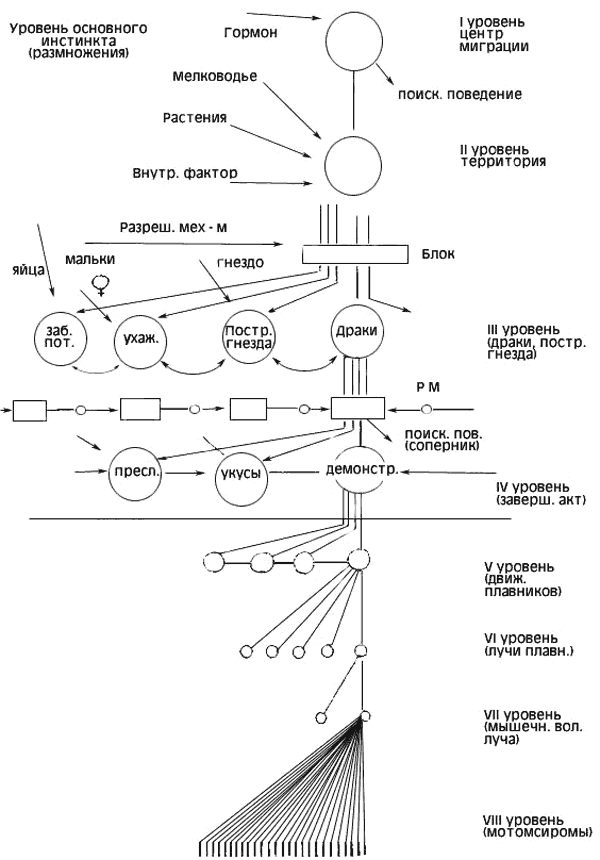
Интерпретируя подобные явления, Тинберген выдвинул гипотезу об иерархии центров, управляющих отдельными поведенческими реакциями (рис.2). Согласно Тинбергену, инстинкт представляет собой завершенную иерархическую организацию поведенческих актов, реагирующую на определенный раздражитель четко координированным комплексом действий. Нужно отметить, что понятие «центр» у Тинбергена является главным образом функциональным и не может быть полностью отождествлено с анатомическим понятием нервного центра, хотя Тинберген и ссылается на известные опыты Хесса. В этих опытах при электрораздражении промежуточного мозга кошек наблюдалось развитие некоторых целостных форм поведения (драк, угроз, сна, пищевого, поведения).
Согласно представлениям Тинбергена, изменение возбудимости центров под влиянием внешних и внутренних воздействий происходит в определенной последовательности. Сначала повышается возбудимость «центра» поисковой фазы поведения, и голодное животное начинает поиск пищи. Когда пища будет найдена, произойдет «разрядка» центра, стоящего на более низком уровне иерархии, и контролирующего осуществления завершающего акта (поедание пищи).
Схему иерархии центров, управляющих поведением самца колюшки в период размножения, Тинберген представляет следующим образом.
Высший центр репродуктивного поведения самца активизируется увеличенной длиной дня, гормональными и другими факторами. Импульсы из этого центра снимают блок с центра поискового поведения. Разрядка этого центра выражается в поисках условий для постройки гнезда. Когда такие условия (подходящая территория, температура, необходимый грунт, мелководье, растительность) найдены, происходит разрядка центров следующего уровня иерархии и благодаря этому становится возможной постройка гнезда.
Если на территорию данного самца проникает соперник, то возбудимость центра агрессивного поведения повышается. Результат этого центра агрессивного поведения — преследования и драки с самцом-соперником. Наконец, при появлении самки повышается возбудимость центра полового поведения, и начинается ухаживание за самкой, представляющее собой комплекс фиксированных действий.
В дальнейшем вопросы иерархической организации поведения изучал Хайнд (1975). Он показал, что хотя в принципе комплекс фиксированных действий большой синицы можно расположить в иерархическую схему, однако не всегда удается сделать это полностью, так как некоторые из движений характерны для двух и более видов инстинктов. Иногда эти движения являются завершающими актами, а иногда — просто средством создания условий, в которых можно осуществить завершающее действие.
У молодых животных иерархия поведения часто еще не сформирована. У птенцов, например, сначала появляются на первый взгляд бессмысленные изолированные двигательные акты, и только позже они интегрируются в сложный функциональный комплекс движений, связанных с полетом.
Расчленение иерархии поведения на элементы часто можно наблюдать во время игры, когда отдельные поведенческие акты, связанные с различными функциями, свободно комбинируются в сочетания не характерных для нормального поведения.
Существенно, что модель Тинбергена предусматривает возможность взаимодействия между «центрами» различных видов поведения. Дело в том, что случаи, когда животное в каждый данный момент занято каким-то одним видом деятельности, являются скорее исключением, чем правилом. Обычно одни виды активности сменяют другие. Наиболее простой пример такого взаимодействия — подавление одних центров другими. Например, если у самца чайки во время ухаживания за самками усиливается голод, то он может прекратить брачные демонстрации и отправиться на поиски корма. В данном случае поведение определяется не присутствием внешнего раздражителя, а соответствующим внутренним побуждением.
Конфликтное поведение
Как особый случай проявления взаимодействия «центров» можно рассматривать так называемое конфликтное поведение, когда у животного наблюдается одновременно несколько тенденций к различным (часто противоположным) типам поведения. Одним из примеров конфликтного поведения — поведение самцов территориальных видов, описанное Тинбергеном в результате наблюдений за трехиглой колюшкой и за различными видами чаек.
Например, если самец А вторгается на территорию самца В, то последний нападает на него и преследует, а самец А спасается бегством. То же самое произойдет, если самец Б вторгнется на территорию самца А. Если же столкновение произойдет на границе этих двух территорий, то поведение обоих самцов будет выглядеть иначе: у обоих самцов элементы реакций нападения и бегства будут чередоваться. Причем элементы нападения будут выражены тем сильнее, чем ближе самец к центру своей территории. Напротив, по мере удаления от центра будут сильнее выражены элементы бегства.
Как показали наблюдения за озерной чайкой Larus redibundus, угрожающее поведение самцов на границе двух территорий включает пять поз, характер и последовательность которых зависят от реакций противника. Каждая из поз отражает определенную степень конфликта между противоположными внутренними побуждениями: агрессивностью — стремлением напасть на противника и страхом — стремлением убежать от него.
Подобный же анализ позволил объяснить и механизм так называемых «замещающих движений» (displacement activity), которые иногда также обнаруживаются у животных в конфликтных ситуациях. Например, в пограничной между двумя участками зоне два самца серебристой чайки (Larus argentatus), стоящие друг перед другом в угрожающих позах, могут внезапно начать чистить перья, белые гуси (Ansercoerulescens) на земле совершают те же движения, что и при купании, серые гуси (Anser anser) в этих ситуациях отряхиваются, а петухи клюют траву и все, что находится поблизости. Эти реакции являются, как выяснилось, врожденными, так как проявляются без соответствующего индивидуального опыта.
Рис. 2. Схема организации инстинкта по Тинбергену
В других случаях конфликт страха и агрессивности приводит к тому, что животное нападает не на противника, а на более слабую особь (как это наблюдал Лоренц у серых гусей), или даже на неодушевленный предмет (чайки при этом клюют листья или землю). Такая «переадресованная» активность, как и «замещающие» действия, проявляется в тех случаях, когда агрессивность и страх уравновешиваются, уступая место другим видам активности, не связанным непосредственно с данной ситуацией.
Таким образом; иерархическая теория инстинктов Тинбергена может объяснить вышеперечисленные явления, и поведение в ситуации конфликта, и замещающие действия, и переадресованную активность.
Работы, начатые Тинбергеном и его сотрудниками, впоследствии, были продолжены и расширены. Накопленный огромный фактический материал (см. например, Хайнд, 1975) показал плодотворность такого подхода и позволил проанализировать многие виды демонстративного поведения. Результаты этих исследований частично соответствовали основным положениям схемы Тинбергена, частично требовали ее усовершенствования. Они как бы продемонстрировали пределы ее применимости и наметили направления ее дальнейшего развития.
Разработка проблем эволюции поведения
Изучение эволюционных аспектов поведения составило одно из основных и наиболее важных направлений в работе Тинбергена и его учеников. Параллельно с Лоренцем они развивали выдвинутые Дарвином идеи об эволюции поведения, последовательно применяя введенный им сравнительный метод и распространяя его на более широкий круг явлений.
Основные работы этологов в этом направлении условно можно подразделить на две основные группы:
1. Выяснение таксономического значения поведенческих признаков. (Работы школы Лоренца).
2. Исследование адаптивного значения поведенческих признаков и роли естественного отбора в их формировании.
Уже согласно определению, поведение является формой приспособления организма к внешней среде. В более узком смысле под адаптивностью поведения понимают те преимущества, которые данный поведенческий признак дает виду в его борьбе за существование. Именно такой анализ и стоял в центре внимания Тинбергена и его учеников. Тинберген писал, что задача этолога состоит не в том, чтобы просто описывать те или иные формы поведения, а в том, чтобы понять, как именно они способствуют борьбе животного за существование.
Свидетельство адаптивности поведения — высокая специализация разных сторон поведенческого акта. О ней говорит прежде всего наличие видоспецифических сложнокоординированных двигательных актов. К ним относятся разнообразные ритуалы, характерные для общественного поведения, движения, связанные с поимкой добычи, строительством гнезд и т. п. реакции, относимые обычно к категории «завершающих актов».
Сенсорные механизмы, обеспечивающие срабатывание реакций типа завершающих актов, также тонко приспособлены к потребностям вида. Завершающий акт выполняется только избирательно в ответ на специфические ключевые раздражители или релизеры. Совершенствование системы сенсорного контроля происходит в двух направлениях. С одной стороны, происходит специализация органов чувств животных, связанных с восприятием ключевых стимулов. Показано, например, что у некоторых животных нейроны слухового анализатора обладают избирательной чувствительностью к специфичным для данного вида звуковым сигналам. Так, например, у пуэрториканкской древесной лягушки Eleutherodcoqulus крик самца состоит из двух слогов — «ко» и «ки». Наблюдения показали, что первый из них «адресован» самцам, являясь, видимо, видоспецифическим территориальным сигналом, второй — самкам. Слияние двух сигналов в один не препятствует существованию раздельных систем их восприятия, напоминающих о соответствии «ключа» «замку». Оказалось, что волоски базиллярной мембраны внутреннего уха самцов и самок этого вида настроены соответственно на разные тоны — либо на восприятие первого, либо на восприятие второго из слогов, составляющих крик самца. С другой стороны, в ряде случаев происходит усовершенствование самих систем, продуцирующих «ключевые раздражители», например звукоиздающего аппарата, а также дифференцированность самих сигналов.
Показано также, что во многих случаях характер онтогенетического развития поведенческого признака также оказывается адаптивным. Там, где поведенческий акт должен быть совершен уже в раннем возрасте (например, взлет бабочки или сосательные движения млекопитающих), жизненно важное значение имеет его «врожденность», и его развитие контролируется главным образом генетическими факторами. В других случаях адаптивность поведения в онтогенезе явно достигается путем обучения методом проб и ошибок. Например, многие птицы обладают врожденной способностью клевать мелкие предметы и поэтому сначала клюют, что попало, и лишь постепенно выучиваются отличать съедобные предметы от несъедобных. Такой тип организации пищевого поведения создает условия для гибкого приспособления к конкретным условиям жизни, когда вид и характер пищи могут варьировать в широких пределах.
Даже такая универсальная характеристика всего живого, как способность к обучению, имеет свою видовую специфику, обеспечивающую наилучшую приспособленность вида, и зачастую представляет собой высокоизбирательный тип взаимодействия с окружающей средой. Например, рабочие пчелы гораздо лучше других насекомых выучивают расположение цветов, на которых они кормятся. Обучение птиц пению происходит, как уже указывалось, в определенные «чувствительные» периоды онтогенеза, причем показано, что птицы предрасположены к усвоению песни своего вида (подробнее см., например, Меннинг, 1982).
В рассмотренных случаях приспособительное значение признаков поведения выступает со всей очевидностью, однако проследить путь их эволюционного развития трудно, или даже невозможно. Особенность работ Тинбергена и его учеников состоит в том, что они, с одной стороны, внесли большой вклад в исследование функционального и приспособительного значения ряда поведенческих признаков, с другой — выделили признаки, на примере которых оказалось возможным показать экспериментально формирующее и стабилизирующее влияние естественного отбора на поведение.
Успехи в выяснения адаптивного значения поведенческих признаков, достигнутые школой Тинбергена, в значительной степени связаны не только с тем, что он четко сформулировал задачу исследований, но и разработал экспериментальные подходы к ее решению. Основным материалом для наблюдений и последующих обобщений послужило поведение нескольких видов морских птиц — чаек, близких в систематическом отношении (одно семейство), но различающихся по ряду особенностей среды обитания, образа жизни, питания и т. п. Основное внимание в этих исследованиях было обращено на изучение поведенческих адаптации, связанных с защитой от хищников, необходимость которой, как выяснилось, накладывает отпечаток практически на все стороны поведения вида.
Необходимый этап в экспериментальном исследовании приспособительного значения акта поведения — его точное описание и выяснение функции, которую он выполняет.
Функциональный анализ признаков поведения
Можно привести много примеров, когда функция поведенческого акта очевидна и не требует специальных исследований. Однако во многих случаях, требуются кропотливые исследования, чтобы выяснить например, функцию отдельных компонентов сложного поведенческого акта. Так, поведение птиц при насиживании имеет вполне очевидную общую функцию. Между тем, для определения конкретной функции отдельных сторон этого поведения, закономерностей их течения во времени и лежащих в их основе регуляторных механизмов потребовались годы интенсивных исследований на птицах разных видов.
В ряде случаев, несмотря на достаточную изученность поведения остаются реакции, функцию которых выявить не удается. Неясно например, почему водные насекомые, прежде чем взлететь, тщательно вытирают поверхность глаз. Не менее загадочной остается описанная у некоторых видов птиц привычка принимать «муравьиные ванны», когда птицы не просто купаются в муравейнике, как в пыли, но могут специально засовывать муравьев себе под перья.
Во многих случаях представление оказалось бы очевидной функции акта поведения после специального анализа совершенно менялось. Так, например, долгое время считали, что специфическая вибрация крыльев у медоносной пчелы служит акустическим сигналом. Однако в настоящее время показано, что его основная функция — вентиляция улья, испарение лишней влаги и охлаждение.
Если функция поведенческого акта не ясна, основой для гипотез служит обычно: сравнительное описание, т. е. сопоставление поведения максимального числа возможных видов, как близкородственных, так и далеких в таксономическом отношении; выявление факторов среды, контролирующих этот акт поведения.
Следует отметить, что экспериментальные исследования функции и адаптивного значения поведенческих признаков при работе в естественных условиях затруднены многочисленными неконтролируемыми и зачастую не поддающимися учету факторами — «шумами». Наряду с этим более строгие лабораторные условия, столь необходимые для физиологических исследований, могут помешать исследователю связать наблюдаемые им факты поведения с успехом выживания вида в естественных условиях.
Сравнительный анализ признаков поведения
При сравнительном изучении признаков поведения этологи анализировали как различие, так и сходство между видами разной степени родства. Благодаря этому удалось выявить наличие адаптивной радиации и конвергенции признаков поведения, т. е. те же закономерности, которые были описаны при изучении эволюции структурно-морфологических, признаков.
Адаптивная радиация.
Адаптивная радиация— это появление разнообразных форм, имеющих общего предка, но видоизменившихся применительно к разнообразным условиям обитания. Адаптивная радиация дает возможность использовать новый источники пищи и избегать врагов.
Классический пример адаптивной радиации — необычайное разнообразие сумчатых в Австралии. Среди них появились травоядные формы — кенгуру, роющие — сумчатые кроты, формы, напоминающие белок и летяг, а также кроликов, волков и других животных.
Примеры адаптивной радиации обнаружены этологами и в поведении, в частности, в общественном поведении животных. На примере многих млекопитающих и птиц показано, что в пределах одного семейства можно наблюдать значительное разнообразие типов группировок, включая как крайние формы, так и переходы между ними. Так, в пределах семейства вороновых можно встретить и виды, ведущие общественный образ жизни в течение всего года (грачи, галки), виды чисто одиночные (кукши), виды живущие в сообществе лишь часть года.
Явление адаптивной радиация поведенческих признаков было описано Тинбергеном и его учениками у чаек. Оказалось, что в зависимости от условий гнездования близкие виды могут существенно различаться по поведению. Например, виды, гнездящиеся на земле, постоянно подвержены опасности нападения как наземных, так и воздушных хищников, поэтому они очень пугливы и поднимают тревогу при появлении рядом с колонией любого живого существа. В отличие от них моевки, которые гнездятся на крутых утесах, совершенно не пугливы и легко терпят около своих гнезд человека, если тому удастся до них добраться. Гнездование моевок на утесах накладывав отпечаток на самые разные стороны их поведения, существенно отличая их от многих других близких видов. Например, отсутствие пугливости по отношению к птицам и животным других видов сочетается у них с гораздо более высокой, чем у других видов чаек, внутривидовой агрессивностью, что обусловлено дефицитом мест, подходящих для строительства гнезд.
Адаптивная радиация признаков общественного поведения происходит, как выяснилось, параллельно в разных семействах, причем, в настоящее время удается выделить те конкретные факторы среды (источники пищи, наличие укрытий, угроза со стороны хищников), которые обусловили появление того или иного типа группировки.
Конвергенция.
Сравнение далеких в систематическом отношении видов — тоже плодотворный метод изучения приспособительных свойств поведения. В ряде случаев оказывается, что организмы, не находящиеся в близком родстве, попадая в сходные условия и эволюционируя под влиянием сходных факторов отбора, в конце концов приобретают сходство. Классический пример конвергенции — водные животные — киты и акулы, относящиеся к разным классам (млекопитающие и рыбы), но сходные по ряду признаков строения тела и образа жизни.
Подобное явление обнаружилось и при сопоставлении поведения далеких в таксономическом отношении видов. Например, разные по происхождению животные, как позвоночные, так и беспозвоночные, которые для защиты от хищников, выработали следующие типы приспособительных маскировочных реакций:
— Сохранение неподвижности при появлении хищника (затаивание);
— Выбор субстрата, по цвету совпадающего с их окраской;
— Выбор позы, способствующей маскировке (создание скрадывающей противотени, имитация различных несъедобных предметов и т. п.;
— Рассредоточение в пространстве несмотря на изобилие пищи.
Один из примеров конвергентного сходства в поведении далеких по происхождению групп животных — это обнаруженная школой Тинбергена синхронизация сроков выведения потомства. При изучении разных видов чаек было установлено, что одновременное выведение потомства — один из способов, обеспечивающих оптимальные условия его охраны от хищников. Показано, что максимальный процент гибели яиц и птенцов наблюдается в выводках, появившихся значительно раньше или значительно позже основной массы, так что чайки-родители оказываются слишком малочисленны, чтобы обеспечить эффективную защиту от врагов на земле и в воздухе. Сходное приспособление существует, по-видимому, и у других животных. Экспериментально установлено, что у антилоп-гну синхронизация сроков размножения обусловлена тем же обстоятельством.
Обнаружение явлений адаптивной радиации и конвергенции поведенческих признаков свидетельствует об общности основных путей эволюции как морфологических, так и поведенческих признаков.
Оценка средовых факторов. В качестве классического примера экспериментального исследования функции и адаптивного значения отдельного поведенческого акта, которое сочетает все вышеуказанные особенности подхода этологов к данной проблеме, можно привести реакции удаления скорлупы у обыкновенной чайки.
Разные виды чаек по-разному поступают со скорлупой яиц после вылупления птенцов. Некоторые не обращают на нее внимания, другие же, в частности, обыкновенная чайка, собирают и уносят подальше от гнезда. Факт этот был известен орнитологам давно, но ему не придавали значения. Однако сам факт наличия этой формы поведения у одних видов и отсутствия у других заставил этологов предположить, что она имеет какое-то значение для выживания вида. Сопоставление биологии этих видов выявило достаточно четкую зависимость: Скорлупу убирают те виды, птенцы которых еще долгое время остаются в гнезде. Если не выводки вскоре после вылупления покидают гнездо, то удаления скорлупы не происходит. Следовательно, эта на первый взгляд незначительная поведенческая реакция имеет определенное адаптивное значение и приносит виду какую-то пользу.
Дальнейший анализ показал, что среди чаек и крачек убирают скорлупу особи тех видов, птенцы которых имеют покровительственную окраску, а не убирают у тех видов, которые имеют достаточно заметных птенцов. Поскольку скорлупа бросается в глаза белизной своей внутренней поверхности, логично было предположить, что после ее удаления гнездо становится менее заметным.
Для проверки этого предположения в колонию чаек исследователи помещали по-разному окрашенные яйца. Выяснилось, что белые яйца. хищники похищали гораздо чаще, чем яйца естественной для чаек окраски. Следовательно, характерная для яиц чаек окраска является маскировочной и имеет существенное адаптивное значение. Однако, как показал Тинберген, покровительственные свойства естественной окраски яиц существенно ослабевают, если рядом положить яичную скорлупу — в таких гнездах яйца гораздо чаще становились добычей хищников. Таким образом, имеются все основания полагать, что удаление яичной скорлупы адаптивно как мера охраны гнезда от хищников.
Однако удаление скорлупок из гнезда происходит не сразу. У обыкновенной чайки, например, они остаются в гнезде 1-2 ч после вылупления птенцов, тогда как у других видов гнездо очищается гораздо быстрее. Оказалось, что эта особенность поведения обыкновенной чайки целесообразна потому, что именно у этого вида взрослые соседи иногда поедают необсохших птенцов, а на обсохших они уже не нападают.
Тинберген указывал, что подобные исследования важны в нескольких отношениях:
1. Способствуют выявлению функции исследуемого поведенческого акта (в данном случае — маскировка гнезда от врагов, нападающих с воздуха).
2. Они экспериментально показывают наиболее вероятные пути формирования данного поведенческого акта в процессе естественного отбора — в данном случае более высокий шанс выжить имело потомство птиц, удалявших скорлупу.
3. Такие исследования позволяют выявить и некоторые дополнительные факторы, влиявшие, вероятно, на формирование этой реакции в процессе естественного отбора — из птиц, уносивших скорлупу, дополнительные шансы на выживание потомства получали те, кто делал это в оптимальный период времени.
Тем самым было показано, что отдельные признаки поведения представляют особой компромиссный результат многочисленных и зачастую разнонаправленных давлений отбора.
Адаптивность и механизмы поведения
Анализ приспособительного значения поведенческих реакций не исключал, а скорее способствовал исследованию, некоторых сторон механизма этих реакций. В частности, при изучении удаления чайками скорлупы наряду с выяснением общей функции этой реакции были изучены стимулы, которые ее вызывают. В результате использования различных моделей было показано, что чайки отличают скорлупу от целых яиц по наличию тонкого контура, тогда как степень изломанности значения не имеет.
Другой вопрос, связанный с первым, — благодаря каким признакам чайки не удаляют скорлупу, пока в ней еще остаются птенцы? Эксперименты показали, что в данном случае решающим признаком будет вес — птицы уносят только легкую скорлупу.
Эксперименты с моделями яиц разного цвета показали, что белую или характерную для чаек крапчатую скорлупу птицы удаляют охотнее, чем скорлупу других цветов. Реже всего птицы удаляли зеленые модели, причем было установлено, что это обстоятельство не связано с дефектом зрительного восприятия. Затормаживание реакции на зеленые раздражители обнаруживает еще раз высокую адаптивность этой формы поведения. Поскольку такую реакцию вызывают предметы с тонким контуром независимо от их формы, то птицы могли реагировать на растения, окружающие гнездо, при этом они часто покидали бы его, подвергая потомство дополнительному риску.
Таким образом, в данном случае исследование механизмов реакции привело к обогащению представлений о ее адаптивности.
Другой пример подобного комплексного подхода — работа ученицы Тинбергена Э. Каллен на моевках. Как уже упоминалось, эти птицы отличаются от других чаек тем, что гнездятся на отвесных скалах, используя для гнезд узкие карнизы. Такой способ гнездования обеспечивает им большую, чем у других чаек безопасность. Обычные наземные враги чаек — лисицы — практически не в состоянии карабкаться по таким уступам, да и для воздушных хищников приземление на карнизы очень затруднено. Эта особенность биологии вида накладывает отпечаток на многие стороны поведения и взрослых птиц, и молодняка. В частности, молодые птицы, практически не падают с карнизов, где расположены гнезда. Анализ показал, что это достигается за счет следующих реакций, свойственных птенцам моевок сразу после вылупления:
1) за счет стремления теснее контактировать с родителями, для которых, в свою очередь, характерно стремление больше времен находиться в гнезде;
2) благодаря стремлению находиться около стены утеса;
3) за счет появления, уже с момента вылупления, боязни обрыва.
Как упоминалось, отсутствие необходимости постоянной обороны от хищников привело к тому, что моевки почти никогда не издают крик тревоги и у них нет системы коллективной защиты, столь совершенной в колониях других видов чаек. Моевки совершенно не боятся человека, но очень агрессивны по отношению друг к другу, так как мест, пригодных для гнездования, очень мало, и за обладание ими птицам постоянно приходится вести борьбу. Характер агрессивных реакций моевок тоже отличается от сходных реакций у других видов. Так, для многих гнездящихся на земле видов характерна так называемая «вертикальная» поза угрозы. У серебристой чайки эта поза является выражением конфликта между побуждениями к двум противоположным действиям — стремлению напасть и стремлению убежать. В этой позе чайка вытягивает шею вверх, а клюв опускает, как бы готовясь клевать противника. Все эти движения эффективны для встречи с противником, нападающим сверху. У моевок же птица-соперник может подлететь не только сверху, но и снизу, и сбоку, поэтому «вертикальная» поза для моевок нецелесообразна. И действительно, наблюдения показали, что в поведения этого вида чаек «вертикальной» позы нет, а моевки нападают на противника, находящегося как выше, так и ниже.
Подобные же особенности отмечены практически во всех проявлениях общественного, агрессивного и родительского поведения моевок.
«Несовершенство» адаптивных свойств поведения
Исследуя адаптивные свойства поведения, Тинберген и др. проанализировали вопросы о так называемом «несовершенстве» адаптивных сторон некоторых форм поведения.
Дело в том, что несмотря на высокую специализацию многих форм поведения, при более глубоком их анализе оказывается, что они имеют недостатки и их нельзя считать наилучшим решением задачи приспособления вида. Примеры неполной адаптивности довольно многочисленны. Так, возвращаясь к поведению морских птиц, можно упомянуть, что несмотря на совершенство и разносторонность системы защиты колонии чаек от хищников вороне время от времени удается «прорваться» и схватить яйцо или птенца. Самец олуши, вида, отличающегося высокой агрессивностью, в процессе ухаживания иногда столь свирепо клюет самку, что она в конце концов покидает его. В разбиравшемся выше поведении чаек при удалении скорлупы из гнезда также имеется недостаток или ошибка, несмотря на всю его высокую адаптивность: чайки-родители, унося скорлупу, покидают гнездо и хищник может схватить птенца или яйцо.
Внимательный анализ «ошибок» обычно показывает, что они чаще всего компенсируются какими-то другими преимуществами. Так, агрессивность, проявляемая самцами олуши (и других территориальных видов птиц) по отношению к самкам, — неизбежное следствие их общей высокой агрессивности, необходимой для завоевания и сохранения гнездового участка. Чайкам-родителям, охраняющим колонию, иногда приходится отступать и жертвовать птенцами или яйцами, так как вороны могут быть опасны и для них самих. Следовательно, окончательная общая эффективность каждого акта поведения определяется соотношением отдельных успехов и неудач для группы особей.
Другой причиной недостаточной адаптивности некоторых форм поведения является «инерционность» процесса эволюции поведения (Майр, 1968). Например, подавляющее большинство птиц при питье зачерпывает воду клювом, а затем глотает, запрокинув голову. Этот способ менее эффективен, чем всасывание воды, характерное для голубей, и тем не менее тенденция к переходу к этому более совершенному способу питья у других видов не наблюдается. По-видимому, такая перестройка связана с какими-то отрицательными последствиями для вида, которые и препятствуют ее осуществлению.
Недостаточная адаптивность некоторых актов поведения связана также с резкими изменениями естественных условий обитания животных. В силу медленного течения процесса эволюции животные, не успевают приспособиться к быстрым изменениям среды, вызванным, например, хозяйственной деятельностью человека. Так, массовое осушение болот в Западной Европе заставляет обыкновенную чайку перемещаться на песчаные дюны, где колонии чаек несут большие потери от нападения лисиц. В болотистой местности их обычно защищает вода, но на дюнах они ничем не защищены от набегов хищников.
Еще один факт, давший повод сомнениям относительно эффективности естественного отбора в отношении признаков поведения, выявился при анализе чувствительности к некоторым ключевым раздражителям. Следует отметить, что изложенные в предыдущей главе представления о врожденном разрешающем механизме и ключевых стимулах Лоренц и Тинберген развивали, работая в тесном контакте друг с другом. В частности, в 1937 г. они опубликовали результаты совместного исследования реакции тревоги только что вылупившихся утят и гусят при виде силуэта летящего хищника. Когда модель переворачивали и двигали «хвостом» вперед, подобной реакции не возникало. Предполагали, что у утят выводковых птиц существует врожденная реакция на силуэт летящего хищника, тогда как при движении силуэта, похожего на гуся, подобная реакция не возникает. Хотя работу Лоренца и Тинбергена неоднократно проверяли и пытались повергнуть с помощью различных модификаций условий опыта и использованных для опыта птиц (подробно см. Меннинг, 1982), однако она во многом определила представления этологов о роли ключевых стимулов в срабатывании врожденного разрешающего механизма. Очень характерен для этологических исследований и использованный в этой работе метод применения модели стимулов, с которыми животное сталкивается в естественных условиях. Подобным образом Тинберген и др. изучили ключевые стимулы, управляющие поведением чаек. Выяснилось, в частности, что можно создать модели, гораздо более эффективные для возникновения данной реакции, чем имитируемые ими естественные раздражители. Например, у птенцов обыкновенных чаек вскоре после вылупления появляется реакция выпрашивания корма. Она состоит в том, что птенец клюет красное пятнышко на клюве родителей. Экспериментируя с различными по цвету, форме и контрастности моделями головы или только клюва Тинбергену удалось показать, что голова живой чайки — не лучший вариант стимула, побуждающего птенца выпрашивать корм, и что можно создать «сверхчайку», гораздо более эффективную, чем реально существующая. Эта «сверхчайка» отличается от настоящей по эффективности весьма значительно, так что возникает вопрос, почему естественный отбор не создал лучшего клюва? Как и в разобранных выше случаях причина кроется, вероятно, в многообразии факторов, под влиянием которых сформировался облик птицы. Дело в том, что отбор закрепляет или отсеивает, не отдельные признаки, а целые фенотипы, поэтому признак, полезный в каком-то определенном отношении, может оказаться неэффективным в других отношениях. Так, например, у птенцов обыкновенной чайки, взрослые особи которой имеют в отличие от серебристых чаек чисто красный клюв, более эффективными оказались модели с контрастным пятном на кончике клюва. Другие наблюдения показали, что красный клюв взрослых чаек играет важную роль в системе сигналов враждебного поведения. Таким образом, облик взрослой птицы сложился как некая совокупность признаков, не всегда максимально эффективных, но зато оптимально действующих в самых разнообразных ситуациях.
Следовательно, представление о якобы несовершенном характере некоторых адаптивных свойств поведения в значительной мере есть результат узости подхода и недостаточно полного анализа всех факторов, контролирующих поведение особи в целом. В ряде случаев это кажущееся несовершенство на деле скомпенсировано какими-то другими преимуществами, как в случае отсрочки удаления скорлупы из гнезда. В других случаях это вынужденный побочный результат отбора на жизненно важный признак, как и в случае самцов олуши, чрезмерно агрессивных по отношению к своим самкам. Во всяком случае, как подчеркивал Тинберген, эти нарушения адаптивности ни в коей мере не говорят об отсутствии роли естественного отбора в происхождении признаков поведения, а наоборот, позволяют выявить некоторые подробности и особенности конкретных механизмов его действия.
Многообразие влияний, контролирующих поведение
Школой Тинбергена проведен ряд исследований, показавших, что признаки поведения формируются и контролируются комплексом разнообразных и иногда противоположно направленных давлений отбора, (например, удаление скорлупы из гнезд).
Многообразие факторов отбора, участвующих в формировании поведения, можно проиллюстрировать рассмотрением системы защиты от хищников у колониальных морских птиц. Система эта очень сложна и включает несколько «линий обороны», причем, чем больше ее изучают, тем больше обнаруживается реакций, в той или иной степени связанных с этой функцией.
Необходимость защиты от хищников накладывает отпечаток на все сферы родительского поведения, начиная со сроков откладки яиц, Известно, что подавляющее большинство членов колонии откладывает яйца одновременно. Ранее предполагалось, что эта синхронность связана с изобилием пищи для выкармливания птенцов. Однако исследования Тинбергена показали, что здесь играет роль не количество корма, а то, что при одновременном появлении птенцов родителям удается лучше всего отгонять хищных птиц. За счет этого смертность одновременно выводимых птенцов оказывается ниже, чем у птенцов, выведенных раньше или позже основной массы. Таким образом, за счет повышенной гибели от хищников ранних и поздних выводков происходит отбор по признаку оптимального срока начала сезона размножения.
Само по себе колониальное гнездование чаек — также способ борьбы с хищниками. Оно позволяет чайкам объединенными усилиями отгонять врагов, нападающих с воздуха, и эта мера тем эффективнее, чем больше численность и плотность колонии. По некоторым данным, на периферии колонии, где гнезда расположены более редко, чем в центре колонии, вороны растаскивают гораздо больше птенцов и яиц.
Существует, однако, и другая опасность — наземные хищники. Их нельзя отпугнуть массовой атакой, часто они нападают именно тогда, когда колония мобилизована против врагов, угрожающих с воздуха, а выводки остаются в гнезде без охраны. Поэтому параллельно происходит отбор признаков поведения, необходимых для борьбы с наземными хищниками. Как показали наблюдения, одним из таких признаков является рассредоточение гнезд, достигаемое за счет «территориального» поведения — угроз и драк между самцами, в слишком плотных колониях лисы похищают больше птенцов.
Таким образом, реально существующая плотность гнезд в колониях формируется в результате компромисса между этими двумя противоположными влияниями — отбором на повышение плотности для защиты от воздушных врагов и отбором на снижение плотности для защиты от врагов на земле.