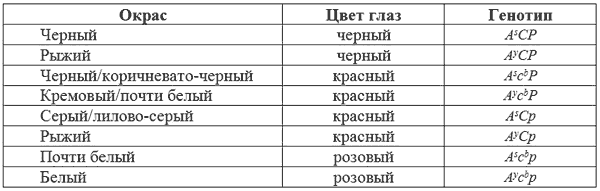
При скрещивании между некоторыми из обнаруженных Персоном и Ашером «альбиносов» были получены щенки с темно-окрашенными шерстью и глазами, что может произойти только, если у родителей за окрас отвечали разные независимые гены, продуцирующие розоватые глаза и осветленный окрас шерсти. Например, кроссы между CCpp (розовоглазое ослабление) а c
bc
bPP (голубоглазое ослабление) дают интенсивно окрашенное потомство (Cc
bPp).
Красноватые глаза не всегда полностью лишены пигмента (в противном случае они были бы розовыми) и радужина у них часто голубоватая.
К сожалению, Персон и Ашер описали всех обнаруженных красноглазых собак как альбиносов, хотя не все они таковыми были. Рассмотрим возможные комбинации взаимодействие локусов А, С и Р.
Гены c
bи p взаимодействуют друг с другом в генотипе c
bc
bpp и продуцируют псевдоальбиносов. При этом, каждый действует так, чтобы удалить пигмент, разрешенный другим. Глаза в этом случае должны быть розовыми, а шерсть почти или совершенно белая. У животных, гомозиготных по b (bb) гены c
bи p могут иметь больший эффект на окрас глаз, чем у имеющих В.
Грифельно-серое ослабление
О гене грифельно-серого окраса известно очень мало. Он наследуется как доминантный и фенотипически сходен с геном ослабления до голубого (d). Этот ген обнаружен лишь однажды у колли (символ Sg).
Ослабление по типу «пуховки»
Это необычное ослабление в значительной степени промежуточно. Щенячья шерсть таких мутантных черных особей сначала серая, потом, к 6-8 месяцам, становится нормальной черной с легким, светловатым оттенком. Ген не оказывает воздействия на желтый пигмент и цвет мочки носа. Ген «пухового ослабления» обнаружен пока только у колли (символ pp) (Лунд и др., 1970). Как действует и как наследуется этот ген неизвестно.
Мраморный окрас (мерль)
Мерль — название, данное для обозначения окраса, при котором наблюдается неровное прокрашивание в виде более темных и более светлых участков одного и того же цвета. Такой окрас встречается у шелти, колли, такс, мраморных догов (арлекинов). Окрас мерль получается в случае гетерозиготного состояния гена М, который доминантен по отношению к исходному т. Светлые участки получаются при смешении нормальных и осветленных волос, причем, это более заметно на черном фоне, чем на рыжем. В гомозиготном состоянии ген мерль дает сплошной белый окрас, частично или полностью голубую радужину глаза, часто редуцированное глазное яблоко, частичную или полную стерильность.
Генотипы наиболее распространенных мерлевых окрасов следующие:
Окрас—
Генотип
Голубой мерль — A
s-Mm
Голубой/биколорный мерль — a
ta
tMm
Рыжий/соболиный мерль — A
y-Mm
При «голубом» или пятнистом мерле районы нормальной и голубой окраски резко различаются. В типичном случае — это голубая собака с черными пятнами неправильной формы, разбросанными на голубом фоне. Сходный эффект можно видеть у черно-подпалого мерля, в большей степени на черпаке и в меньшей — на животе. Ген М — больше влияет на черный/коричневый пигмент, нежели на желтый, поэтому, у чисто рыжих собак мерлевая пятнистость не так явно выражена. Замечено, что ген М усиливает белую пятнистость.
Из-за того, что ген мерль в гомозиготном состоянии несет ряд нежелательных признаков, не следует скрещивать гетерозигот, а тем более гомозигот между собой. Логичнее получать мраморных собак, скрещивая мраморных и нормально окрашенных особей. В этом случае получатся как мраморные, так и нормальные щенки.
Возможно, что существует более, чем один аллель гена мерль. Это следует из различий в фенотипическом проявлении и результатов скрещивания (Schaible и Brumuiugh, 1976). Одним из них можно считать ген M
h, обуславливающий мраморный окрас дога (арлекин). Этот ген продуцирует белые пятна в гетерозиготном состоянии (M
hm)- Гетерозиготы M
hM сходны с ММ по фенотипическим проявлениям (белый окрас, частичная глухота, микрофтальмия). Признаки M
hM
h, возможно, также сходны с ММ или даже более выражены.
CN — ослабление
Этот ген ослабляет как черный пигмент, так и желтый: черный до тускло-серого, а желтый до бежевого или почти белого. Соболиный окрас трансформируется в серебристо серый или почти белый в соответствии со степенью затененности. Мочка носа окрашена в светло-рыжевато-коричневый цвет, что заметно отличает действие гена cn от действия других генов ослабления окраса. Волос CN-щенков имеет более тонкое строение и может даже казаться слегка волнистым. Этот эффект исчезает у взрослых. Ген «CN — ослабления» наследуется рецессивно (символ cn) и является полулетальным, вызывая дефицит нейтрофилов крови, что серьезно влияет на способность индивидуума противостоять бактериальной инфекции. Большая часть щенков, несущих cncn погибает в течении нескольких первых месяцев жизни. «CN — ослабление» пока известно только для колли.
Прогрессирующее поседение
Литтл (1957) предположил, что прогрессирующее поседение, или посерение, которое наблюдается у некоторых пород собак, происходит благодаря действию полудоминантного гена G. Автор не приводит экспериментальных данных по скрещиванию для подтверждения своего предположения, однако, Витни (1952, 1958) опубликовал результаты, которые указывают на то, что прогрессирующее поседение детерминировано одним геном. Сначала он обозначил этот ген как si, потом согласился с термином Литтля (G). Характер наследования гена (рецессивный, полудоминантный или доминантный) до сих пор еще не ясен окончательно.
Термин «поседение» употребляется в генетике млекопитающих в случае, когда надо отметить появление значительного количества белых волос на теле. В этом случае количество темных волос превосходит количество белых и животное кажется темным, испещренным белыми волосками. Если количество белых волос больше, чем темных, то такой окрас называют чалым. Непонятно пока, то ли это две отдельные генетические формы, та ли различные степени экспрессии одного гена.
Поседение также классифицируется, как постоянное и прогрессивное. В первом случае оно возникает на определенной стадии развития и остается неизменным с возрастом. Прогрессивное поседение возникает постепенно на протяжении значительного периода или всей жизни. Поседение по второму типу связано с геном G.
Щенки, имеющие ген G рождаются черными (или интенсивно окрашенными), потом сереют (светлеют). Гетерозиготы Gg могут изменяться до темно-серо-голубого. В некоторых случаях это происходит достаточно быстро, но, в основном, животные седеют на протяжении всей жизни. Сходные изменения происходят с гомозиготами, но более резко и глубоко.
Поседение начинается в возрасте нескольких недель и со временем окрас становится серо-голубым. Причем, степень выраженности варьирует как от породы, так и от индивидуума. Поседение может происходить равномерно по всей поверхности туловища, а может и частично. Так, например, у бедлингтона верх головы и часть плеч могут стать почти белыми.
В комбинациях с другими аллелями Gg дает слабое осветление окраса, тогда как GG- резкую редукцию пигмента.
Осветление окраса за счет генотипа GG означает, что ген G если не полностью, то частично эпистатичен по отношению к другим генам окраса. Приведем ниже генотипы, представляющие наибольший интерес:
Окрас—
Генотип
Серебристый — A
s-B-D-E-GG
Шоколадно-серебристый — A
s-bbD-E-GG
Голубо-серебристый — A
s-B-ddE-GG
Кремово-серебристый — A
s-B-D-eeGG
Светло кремовый, серебристый — A
s-B-ddeeGG
Животные имеющие вышеперечисленные окрасы являются серебристыми или чалыми, у которых белые или слабопрокрашенные волосы перемежаются с окрашенными. При этом, если на тела много окрашенных волос, то цвет их легко фенотипически идентифицировать, но, если их мало, определить окрас можно лишь по общему впечатлению. Так, все bb можно идентифицировать по наличию коричневой мочки носа и осветленных глаз. Шерсть может быть с легким коричневатым оттенком. Таким же образом ee-серебристые животные могут иметь бледно-желтый или кремовый оттенок. Все фенотипы можно легко распознать до начала поседения. Следует отметить, что A
s-B-ddE-GG должны иметь голубой окрас даже до начала поседения благодаря наличию гена d, а не G. Экспрессия гена G может быть не так заметна благодаря аффекту ослабления за счет гена d. Любая порода, где какие-то особи рождаются интенсивно окрашенными, а затем постепенно бледнеют до отчетливо более светлого окраса, имеет в своем генофонде ген G. Очевидно, что он имеется у таких пород, как бедлингтон, керриблютерьер, бобтейл, пудель. Литтл (1957) предложил, что ген G может присутствовать у кернтерьера, денди-динмонт-терьера, скайтерьера и йоркширского терьера. Можно также предположить его наличие у австралийского терьера и австралийского шелкового терьера.
Белая пятнистость
Наличие белого узора на туловище характерно для многих пород собак. Более того, у многих из них характер расположения белых отметин является чуть ли не основным признаком. Теоретически, белые участки рассматриваются как белые пятна, не взирая на количество белого. Раньше этот термин употреблялся только, когда количество белого лимитировано в виде пятен на темном фоне. Теперь термин «белая пятнистость» употребляется более широко и включает также случаи, когда белый преобладает или же собака почти вся белая. При этом белое поле можно рассматривать как одно большое белое пятно.
Распределение и распространение белых отметин следует относительно правильной прогрессии. Так, если расположить пятнистых собак по мере убывания прокрашенных участков, то выявляется следующая закономерность (см. рис. на стр. 87). Сначала белые отметины появляются на груди, лапах, морде и кончике хвоста. По мере увеличения площади белый распространяется на всю грудь, живот, конечности. Долее — на плечи и холку, затем бока и верхнюю часть спины. Постепенно, на теле остаются немногочисленные пигментированные пятна, уменьшающиеся в размерах. На этой стадии также наблюдается закономерность в расположении оставшихся окрашенных пятен. Чаще всего они остаются на боках, голове, особенно вокруг ушей и в основании хвоста. На рисунке схематично отображены описанные процессы.
Причиной такой общей закономерности развития белой пятнистости состоит в отсутствии пигмент-образующих клеток в коже раннего эмбриона при корнях волос. Эти клетки образуются в определенных местах, что соответствует приблизительно участкам на голове и вдоль спины у развивающихся эмбрионов. Если что-то препятствует образованию достаточного количества первичных точек пигментации или скорость распространения пигмент-образующих клеток по туловищу тормозится в ходе развития эмбриона, то щенки рождаются с белыми отметинами. Хотя структура кожи остается нормальной и на ней вырастают нормальные волосы, они будут бесцветными (т.е. белыми для человеческого глаза), так как пигмент-продуцирующие клетки не успели распространиться в эти участки. Так как эмбриональное развитие — это строго детерминированный процесс, то для этих клеток возможность своевременно занять свою позицию так же строго ограничена во времени.
Следует, однако, отметить, что, несмотря на общие тенденции в увеличении количества белого, существуют значительные вариации. Особенно это касается собак с преобладанием белого. У них пятна могут возникать в любом месте головы, плеч, боков и, как правило, не обязательно симметрично. Напротив, положение белой проточины на морде и отметин, образующих воротник (так называемая воротниковая степень пятнистости) относительно постоянно и служит отличительной чертой таких пород, как бостон-терьер, колли и сенбернар.
Животные, у которых на белом фоне разбросаны пигментированные пятна, называются пегими. Некоторые из этих вариаций обусловлены действием полигенов, модифицирующих экспрессию (т.е. количество белого) основных аллелей пятнистости, а некоторые — нарушением в ходе эмбрионального развития.
Показано, что, если в породе предпочтителен какой-то определенный характер пятнистости, то количество других вариаций значительно уменьшается, но не исчезает совсем.
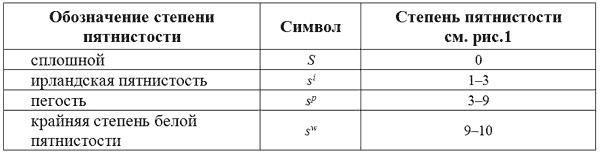
Литтл (1957) выдвинул гипотезу, что разнообразие распределения пятен на туловище можно объяснить существованием трех мутантных аллелей гена S, определяющего сплошной (небелый) окрас.
Аллель ирландской пятнистости продуцирует наименьшую степень пятнистости. Белые отметины варьируют от пятен начиная с морды, груди, плеч, головы, ног и кончая узким воротником на шее. Типичным примером экспрессии вариабельности ирландской пятнистости s
is
iпо Литтлю является бассенджи.
Животные, гомозиготные по гену пегости s
ps
p, имеют большее количество белого, чем животные предыдущего типа. Однако, для s
pхарактерно значительно более широкая вариабельность экспрессии. Например, по Литтлю, бигль имеет генотип s
ps
p, но значительная часть белых отметин по характеру распределения лишь ненамного отличается от бассенджи. С «другой стороны фокстерьеры имеют такой же генотип s
ps
p, но в общей своей массе имеют столько белого, что порой их можно рассматривать как белых собак с небольшими пигментированными пятнами.
Аллель крайней степени пятнистости определяет наибольшее количество белого и, в крайнем случае, такие животные полностью белые за исключением небольшого пятна или пятен на крупе или голове. В эту группу также входят темноглазые собаки совсем без пятен. У целого ряда пород такой фенотип является или стандартным окрасом или принятой вариацией.
Доказательством того, что чисто белые темноглазые собаки имеют генотип s
ws
wявляется тот факт, что у таких собак могут иногда появляться щенки с небольшими пятнами в стандартных местах. Поэтому, Литтл предположил, что белые бультерьеры, большие пиренейские собаки, самоеды и салихемтерьеры имеют генотип s
ws
w. Возможно, что аллели пятнистости не полностью доминантны друг к другу, но эти взаимоотношения неустойчивы.
Исходя из многих наблюдений, можно предположить, что ген S доминантен к s
i, но не к s
pили s
w. Гетерозиготы Ss
pи Ss
wдают окрасы по типу ирландской пятнистости. Аллель s
iдоминантен к s
p, но не к s
wГетерозиготы s
is
wдают пегость или окрас, близкий к этому. Взаимоотношения между s
pи s
wпока не ясны из-за недостатка данных. Возможно, что s
ps
wдают белых собак с незначительным количеством окрашенных пятен. Следует помнить, что экспрессия s
pпоказывает огромную вариабельность и серьезно усложняет окончательное выявление взаимоотношений между аллелями этого локуса.
Кроме этого, экспрессия всех мутантных аллелей пятнистости частично контролируется модифицирующими полигенами. В большей мере они определяют независимо от породы степень пятнистости, нежели порядок, местоположение пятен и их конфигурацию.
До сих пор не выяснены механизмы действия модификаторов: то ли они действуют сами по себе, то ли влияют на экспрессию S-аллелей. Почти очевидно, что некоторые из них способны сами по себе продуцировать начальную степень белой пятнистости у гомозиготных SS (генотипически — это генотип сплошного окраса). В некоторых случаях небольшие белые пятна могут возникать как следствие сбоев эмбрионального развития. Однако, сомнительно, чтобы все подобные случаи можно было бы объяснить таким образом. Ситуация усложнена еще и тем, что многие полигены, вовлеченные в этот процесс, экспрессируются также вариабельно. Таким образом, многие SS-животные, имеют незначительные белые отметины за счет действия модификаторов. В любом случае, при постоянном отборе только полностью пигментированных собак, всегда будут случаи появления небольшой части животных с белыми отметинами.
И Витни (1947) и Литтл (1957) отмечали, что черные и красные спаниели отличаются по количеству белого. Так, у большей части черных собак наблюдается небольшое или среднее количество белого, тогда как у рыжих спаниелей белого значительно больше. Можно предположить, что один из наиболее влиятельных модификаторов гена s сцеплен с геном е, или, что пигмент-продуцирующие клетки, содержащие в геноме ее менее способны распространяться по коже сравнительно с E-содержащими. Такие различия не проявляются у полностью окрашенных животных, но становятся очевидными у пятнистых. Берне и Фразер (1966) отметили, что коричневые собаки занимают в этом отношении промежуточное положение. Это подразумевает, что ген b является фактором, также влияющим на развитие пятнистости.
Может показаться странным, что различные гены белой пятнистости являются аллелями одного локуса, так как у многих видов млекопитающих белая пятнистость возникает при мутациях различных локусов. Сходную ситуацию можно было бы предположить и для собак, как отмечает Хатт (1979). Однако, обширные данные, опубликованные Литтлем (1957) полностью согласуются с предположением о существовании только одного локуса с несколькими аллелями. Существование независимого от s-аллелей гена белой пятнистости сомнительно. Вопрос лишь в том, сколько на самом деле аллелей: две или три, так как очень трудно порой различить экспрессию s
iи s
p.
Тиковость (тиковая пятнистость, крапчатость)
У многих пород, таких, как пойнтеры, сеттеры и спаниели, белые участки покрыты многочисленными пигментированными мелкими пятнами. Такое явление называется тиковой пятнистостью и определяется геном Т. Тиковая пятнистость отсутствует при рождении и проявляется в возрасте от нескольких недель жизни щенка до первой линьки. Экспрессия гена Т очень вариабельна: от нескольких небольших пятнышек до такого их изобилия, что белые участки тела кажутся чалыми. У длинношерстных пород тиковость выглядит как чалость благодаря тому, что длинные волосы разных цветов легко перемешиваются. У некоторых пород наблюдается тенденция к усилию тиковости на ногах и животе.
Тиковость является результатом взаимодействия между любой из аллелей белой пятнистости и гена Т. Чаще всего это аллель s
pкак наиболее распространенный и продуцирующий белые участки достаточной протяженности для четкого проявления тиковости. Тиковость не влияет на гены окраса, т.е. цвет крапа соответствует основному окрасу. Генотип особей с тиковой пятнистостью должен быть s
ps
pT-, s
ps
wT- или s
ws
wT-. При этом следует помнить, что количество белого очень вариабельно, и, что тиковость проявляется только на белом фоне. Остальная часть генотипа, определяющего окрас, будет соответствовать цветовой вариации особи. Например, тиковый черный имеет генотип A
s-B-D-E-s
p-T-, а рыжий тиковый A
y-B-D-E-s
p-T-. Черно-подпалые тиковые (a
la
lB-D-E-s
p-T-) имеют черный крап на спине и рыжий на животе.
Раньше считали, что пятнистость по типу далматинов определяется отдельным геном, но Литтл (1957) убедительно показал, что это модификация обычной тиковости. Например, черные далматины имеют генотип A
s-B-D-E-s
ws
wT-. Разведенцы далматинов вывели собак с более крупными, яркими, прокрашенными пятнами, случайно разбросанными по всему телу.
Экспрессия гена тиковой пятнистости Т варьирует от слегка крапчатые собак с несколькими крапивами только на конечностях и животе до сильно крапчатых животных, часто встречающихся у гончих пород (например, гасконская гончая и гриффон). В последнем случае собаку с таким окрасом можно назвать чалой. С другой стороны, варьирует не только количество крапа, но и качество его. Так что тиковость является весьма вариабельным признаком. Является ли это результатом отбора и закрепления в генотипе определенного количества полигенов, управляющих экспрессией гена Т, еще предстоит выяснить. Литтл заметил, что при скрещивании далматинов и собак с окрасом, похожим на далматинский, в первом поколении происходит возврат к обычной крапчатости. Возможно, что существует не один ген тиковости, каждый со своим характерным типом крапчатости. Однако, тот факт, что у собак наблюдается экстраординарная вариабельность экспрессии тиковости сам по себе не может служить этому доказательством.
Маски
Маска — термин, употребляющийся для обозначения черного поля на морде, часто покрывающего переднюю часть «лица» и ушей. Наиболее четко маска видна у А
у-рыжих собак, например, у таких разных пород, как желто-коричневая (оленья) вариация гигантского мастифа и у миниатюрного мопса.
У чепрачных и черно-подпалых собак маска, если она есть, затемняет подпалы, ассоциированные с генами a
saи a
t. В этом случае получаются черномордые чепрачные и черно-подпалые собаки. У первых, ген маски трудно выявить, т.к.
наличие или отсутствие темной морды можно также рассматривать и как вариации чепрачности. Черно-подпалый окрас более стабилен и однообразен, поэтому маска у черноподпалых собак сразу же обнаруживается, благодаря отсутствию подпалов на морде. Естественно, что маску невозможно обнаружить у черных A
s-собак, хотя в геноме соответствующий ген может присутствовать.
Маска может варьировать по степени развития. Так, она может покрывать всю «лицевую» часть головы и уши. Но это не обязательно означает влияние полигенов. О характере наследуемости маски известно очень мало. Литтл (1957) ссылается на случай рождения от рыжей таксы и мопса оленьего окраса с маской всех восьми щенков с черными масками. Это указывает на моногенность и доминантность маски как признака. Условно, ген маски можно обозначить как Ма.
Литтл считает, что существует несколько аллельных генов маски, но не дает четких доказательств. Тигровые животные могут быть как о маской так и без нее, что само по себе достаточно, чтобы отбросить всякие сомнения о моногенности признака. Геи маски, возможно, не экспрессируется у ee-животных, но и это не свидетельствует о наличии серии аллелей. Возможно, что ee-рыжие животные не могут вообще синтезировать черный пигмент в волосе. Отсутствие маски у ee-рыжих собак можно рассматривать как признак, отличающий их от А
у-рыжих животных. Все рыжие собаки с маской имеют ген А
у, тогда как рыжие без маски могут иметь как Ay так и ee, но, скорее всего последний.
Руфизм (рыжина)
Интенсивность желтого пигмента шерсти может варьировать от бледно-кремового до насыщенного рыжего. Некоторые из вариаций можно рассматривать как результат действия основных генов окраса, таких как d или, возможно, c
ch(шиншилла), но не все. Остается еще значительное количество вариаций, которые обязаны действию группы полигенов, известных как руфус-полигены. Причем, эти полигены воздействуют как на A
yтак и на ее. Интенсивность пигментации варьирует в зависимости от особи, линии или породы. Эти же модификаторы также определяют интенсивность пигментации подпалов у чепрачных и черно-подпалых собак, у которых они могут варьировать от бледно-желтых до насыщенно красных. Это поднимает статус данной группы генов о простых модификаторов генов А
уи е до фундаментальной группы генов, определяющих основной составной элемент окраса. Только сплошной черный или сплошной белый окрасы не подчинены влиянию руфус-полигенов.
Действие этих генов в основном направлено на усиление пигментации. Волк имеет тусклый, однообразный окрас и собаководы прошлого, очевидно, отбирали более яркоокрашенных особей, определяя таким образом внешний вид большинства пород. Ильин (1932, 1941) обнаружил, что более бледный окрас доминантен по отношению к более яркому и объяснил это существованием серии аллелей «интенсивности» (символы Int, int
mи int). Аллель Int продуцирует наиболее бледные тона, a int — наиболее яркие, int
mзанимает промежуточное положение. Вполне возможно, что интенсивность желтой пигментации действительно регулируется мажорными генами (а не полигенами), но, чтобы доказать это, у Ильина было слишком мало наблюдений. Скорее всего, в этот процесс вовлечены многочисленные руфус-полигены. Наблюдения Ильина интересны и, возможно, полезны, т.к. потерю яркого рыжего окраса у гибридов между яркими и осветленными собаками наблюдали Njebbes и Wriedt (1927). Бернс и Фразер (1966), а также Виллис (1976) заметили, что насыщенность рыжего окраса может быть легко потеряна и трудно затем восстанавливается. Это означает, что, если есть усиливающие и ослабляющие полигены, то последние должны быть доминантны по отношению к первым. Однако, для этой группы полигенов характерно, что те, которые доминантны в одних скрещиваниях, становятся рецессивны в других, в зависимости от родителей. Т.о. очень трудно выявить какие-либо закономерности, лежащие в основе взаимодействия этих генов.
Вариабельность чепрачности
Экспрессия гена чепрачности настолько вариабельна, что можно предположить наличие более, чем одного аллеля, определяющего этот признак. Например, один аллель дает темно-чепрачный окрас, другой — светло-чепрачный. Вариации чепрачности составляют непрерывный ряд, что можно объяснить скорее влиянием модифицирующих полигенов. Чепрачный окрас часто можно спутать с соболиным, т.к. он имеет тенденцию с возрастом осветляться. Это говорит о генетической близости аллелей a
saи A
y. С другой стороны, когда чепрак распространяется на большую часть тела, окрас становится похожим на черно-подпалый.
В идеале, чепрачными принято называть собак с V-образным темным пятном на спине и боках туловища, простирающимся от хребта до живота. Однако, как распространение, так и интенсивность пигментации этой области может варьировать. При яркой, интенсивной пигментации чепрачный окрас легко идентифицировать, но при более светлой пигментации границы чепрака размыты и наличие его трудно распознать. Фактически, этот окрас стремится иметь сходство с диким волчеобразным. И это является основой для предположения, что ген чепрачности является модификацией исходного гена А. Сходство усиливается, если рыжий подпал осветлен. Строго говоря, вариабельность чепрачности определяется двумя независимыми системами модификаторов. С одной стороны — это модификаторы, управляющие вариабельностью чепрака, его интенсивностью, формой и протяженностью. С другой стороны, окрас подпалов определяется руфус-полигенами, как в случае с чисто рыжими собаками. Причем, они не влияют на экспрессию гена чепрачности.
Интересно было бы предположить, что некоторые фенотипические вариации чепрачного окраса обязаны полигенам, влияющим на экспрессию гена А, которые затеняют рыжий окрас превращая его в соболиный. Такие затеняющие полигены могли бы продуцировать некоторые вариации чепрачного окраса, модифицируя экспрессию гена А. К сожалению, никакими экспериментальными данными по этому вопросу мы не располагаем. Интенсивность пигментации подпалов может также варьировать, причем независимо от интенсивности окраса чепрака. Почти наверняка это происходит благодаря действию независимых руфус-полигенов.
Полигены затенения или соболиного окраса
Эти полигены, судя по названию, вызывает потемнение шерсти. Их наиболее очевидной экспрессией является измелен не обычного А
у-рыжего окраса до соболиного. Большинство, если не все А
у-рыжие особи имеют некоторое количество черно-типированных волосков, как правило, на голове и вдоль спины. Туловище щенка обычно покрыто темными волосами, придавая ему более темный окрас. При взрослении щенка окрас становится все более светлым, пока не будет достигнут окрас взрослого животного.
Когда количество темных волос многочисленно у взрослого животного рыжего окраса, получается соболиный окрас. Степень затененности значительно варьирует, особенно на голове, плечах и спине. Наиболее темные особи могут напоминать осветленных чепрачных. Особенно, если затенение распространяется на бока, напоминая по форме сглаженный чепрак.
Типичный соболиный окрас можно встретить у колли или шотландской овчарки. Показано, что только А
у-рыжий окрас конвертируется в соболиный с помощью полигенов затенения. Предполагается, но еще не доказано, что эти полигены не оказывают сходного воздействия на ген е, который, очевидно, эпистатичен по отношению к данным модификаторам.
Белые собаки
Фенотипически, полностью белые собаки могут быть основой для ряда пород или породных вариаций. В принципе, полностью белые собаки имеют различную генотипическую основу. У многих млекопитающих встречается альбинизм. Этот фенотип полностью лишен Пигмента не только в волосах, но и в радужине глаза, кожи, когтях и др. Такие белые особи красноглазы. Однако, когда собаководы говорят о белых собаках, они имеют в виду темноглазых животных. Таких собак нельзя назвать альбиносами, даже если они имеют голубые глаза.
Одно из наиболее ранних предположений о происхождении чисто белых собак состоит в том, что окрас таких особей есть следствие действия гена крайней степени пятнистости. Литтл постулировал существование аллеля s
w, который и обуславливает чисто белый окрас. Животные с генотипом s
ws
wили полностью белые, или имеют незначительные отметины в области ушей и основании хвоста. Постоянная селекция и удаление особей с отметинами из разведения в прошлом привели к тому, что они составляют небольшую часть популяции. Однако, факт, что такие особи все-таки появляются, является хорошим подтверждением существования генотипа s
ws
w. В результате, Литтл смог предположить, что у бультерьера, большой пиренейской собаки, самоеда и силихэмтерьера имеет место быть генотип s
ws
w.
Можно также предположить, что чисто белый окрас возникает в результате действия гена шиншилла (c
ch), который удаляет почти весь желтый пигмент волос, но не глаз. В результате могут получиться темноглазые белые собаки. Tjebbes и Wriedt (1927) предположили, что такими собаками могут быть самоеды. Возможно, что они ошибались, исходя из более поздней концепции Литтля. Однако, совсем не обязательно, что все самоеды несут s
ws
w, т.к. данные Литтля получены при исследовании американской популяции самоедов, а Tjebbes и Wriedt наблюдали скандинавскую группу.