Прежде, чем приступить к изложению генетики пород собак, необходимо в общих чертах ознакомиться с тем, что известно о генах, определяющих окрасы и структуру шерсти у собак, и их действии. Для этого было бы полезно иметь представление об общих принципах наследственности, с которыми можно ознакомиться в популярных, и специальных изданиях по генетике. В настоящей главе мы обсудим действие генов более детально и систематизировано. В основном использовались общепринятые в генетике млекопитающих символы для обозначения генов и их мутантов.
До сих пор было проведено ничтожно мало контролируемых экспериментов по скрещиванию собак. Такая работа требует значительных расходов и длительного времени. Поэтому, в некоторых случаях использованы результаты случайных наблюдений. Однако, основная масса информации по генетике собак была получена из достоверных источников.
Аллели «Агути»
Ряд наиболее распространенных фенотипов окраса определяются серией аллелей, известной как «агути». Этот термин произошел от названия небольшого грызуна с коричневосерой шерстью, которая прекрасно скрывает его от хищников. У псовых соответствующий окрас называют волче-серым (зонарным). Однако, в этом случае, такой окрас позволяет хищнику незаметно подкрадываться к жертве.
В генетике собак термин «агути» позаимствован из генетики грызунов, где ген «агути» хорошо изучен. Выяснено, что исходный (дикий тип) ген «агути» является родоначальником целой серии аллелей. В настоящее время выявлены следующие аллели:
Фенотипическое проявление—
Символ
Сплошной черный — A
s
Доминантный желтый (рыжий) — A
y
Агути (волче-серый) — A
Чепрачный — a
sa
Черно-подпалый — a
t
Исходный ген А («агути») отвечает за волче-серый окрас диких псовых и большинства, если не всех пород собак, имеющих волчеобразный серый окрас. К таким породам можно отнести серую сибирскую хаски, серого норвежского элькхунда и других собак крайнего Севера. Возможно, что серая немецкая овчарка имеет генотип АА.
В результате мутагенеза ген А дал четыре мутантных аллеля: два доминантных (A
sи A
y) и два рецессивных (a
saи a
t) по отношению к исходному гену А.
Ген A
sотвечает за сплошной черный окрас, характерный для многих пород. Этот окрас варьирует от угольно-черного до буро-чёрного. Последний образуется в результате различного числа остаточных агути-подобных волос. Нечистокровные черные собаки часто имеют бурый оттенок. При ближайшем рассмотрении их шерсти можно выявить некоторое количество зонарных волос наряду с чисто черными. Наиболее простое объяснение этому явлению заключается с том, что ген A
sсам по себе не может индуцировать образование чисто черных волос. Для этого ему требуется наличие достаточного количества модифицирующих полигенов, селекционно отобранных при выведении чистокровных пород. Можно также предположить, что ген A
sне полностью доминантен, тогда генотип A
sA
sопределяет чисто черный окрас, а генотипы A
sA
yили A
sA — бурый. Агути-подобные волосы могут давать также специфический красноватый оттенок, описанный Литтлем (1957), который предположил, что такой отгонок образуется благодаря неполному доминированию A
sнад A
yи a
t. Литтл показал, что у особей A
sA
yкрасноватый оттенок встречается в основном на боках, голове, шее и конечностях, а особи с генотипом A
sa
tимеют красноватый оттенок только там, где должны располагаться подпалы у особей a
ta
t. Такое неполное доминирование вполне возможно. Однако, следует заметить, что статистически это еще не доказано.
Доминантный аллельный ген A
yотвечает за рыжий окрас собаки. Этот ген дает чисто рыжих собак, но при этом обнаруживаются волосы с черными кончиками (т.н. типированные) на голове, плечах, вдоль спины, включая хвост. При наличии значительного количества таких волос можно говорить о соболином окрасе. Поэтому, этот аллель можно назвать также аллелью соболиного или соболе-рыжего окраса. По этой терминологии полностью, рыжие формы можно обозначить как золотисто-соболиные или светло-соболиные. Количество зачерненных волос или степень проявления соболиного окраса варьирует, что обусловлено полигенами, которые наследуются независимо от А. В совокупности, эти полигены известны под названием «затеняющие».
Литтл (1957) предположил, что соболиный окрас проявляется при гетерозиготном генотипе A
ya
t. Тогда, при скрещивании обоих особей соболиного окраса, потомство должно быть рыжим, соболиным и черно-подпалым в соотношении 1:2:1 соответственно. Экспериментально, примерно так и получалось. Однако, и от чисто рыжих собак также получались периодически черно-подпалые щенки, что говорит о гетерозиготности обоих родителей (A
ya
t). Это обстоятельство в значительной степени опровергает идею неполного доминирования A
yнад a
t. Можно предположить, что полигены затенения сами по себе так взаимодействуют с генами a
tи A
y, что в случае гетерозиготы A
ya
tполучается темно-соболиной окрас, а в случае гомозиготы A
yA
y— рыжий окрас со слабым или незначительным затенением. Но это весьма сомнительно.
Ген чепрачности a
saопределяет V-образный характер пигментации шерсти по обеим сторонам туловища, как, например, у эрдельтерьера или бигля. На первый взгляд, фенотипы a
saи a
tсходны, но чепрачные особи имеют больше рыжего, чем черно-подпалые, особенно на морде, плечах, боках и конечностях. Однако, большое распространение черного в чепрачном окрасе может напоминать черно-подпалый. Из двух этих окрасов чепрачный наиболее вариабелен.
В типичном случае черно-подпалого окраса (доберманы, например) черный распространяется на всю верхнюю часть туловища, а рыжий ограничен внутренней поверхностью конечностей, грудными отметинами и нижней поверхностью морды. Два характерных пятна расположены над глазами. Для чепрачного окраса характерны возрастные изменения. Так, чепрачные щенки рождаются очень похожими на черноподпалых, с возрастом они светлеют, рыжие отметины увеличиваются по площади, пока молодые животные не становятся истинно чепрачными. Нельзя не отметить интригующий факт, что соболиные собаки подвержены сходным возрастным изменениям. Это явление настолько всеобщее, что и детеныши волка имеют более темный окрас, чем взрослые животные.
Интересно, что у большинства чистокровных пород собак, за исключением эскимосских и скандинавских, аллель дикого типа А отсутствует. Такое отсутствие столь характерно, что кажется преднамеренным. Возможно, что древние селекционеры стремились закреплять такие цветовые вариации, которые, во-первых, отличали бы домашних собак от диких форм и, во-вторых, служили бы символом одомашнивания собак.
Наиболее очевидно, что самым близким аллелем к гену А является аллель a
sa. В основном А и a
saпроявляются сходно. Так, у нечистокровных животных наблюдаются все переходы от зонарного (волче-серого) до чепрачного окрасов. Более того, Фокс (1978) предположил, что зонарный и чепрачный окрасы обусловлены действием одного и того же гена. Он скрещивал койота и бигля и в последующих двух поколениях получил в общей сложности 16 животных, которые летом были темно-соболиного окраса, а зимой светлели до светло-соболиного. Однако, по рисункам, представленным Фоксом, потомство было скорее зонарным или светло-чепрачным. В любом случае, это наблюдение показательно, но не убедительно.
Литтл (1957) предположил, что чепрачный и черно-подпалый окрасы обусловлены действием одного аллеля, а различия между ними являются результатом работы модифицирующих полигенов. Однако, в 1976 году Виллис убедительно продемонстрировал различия между генами a
saи a
tи доказал доминирование a
saнад a
t. Не очень правдоподобна также и идея о существовании более одного аллеля, кодирующего чепрачный окрас. В этом случае наблюдались бы явные скачкообразные различия, тогда как встречаются постепенные переходы от самых затемненных до самых светлых чепрачных окрасов.
Кажется возможным, что у немецкой овчарки черный окрас кодируется не геном A
s. Так, Виллис описал черную форму, рецессивную по отношению ко всем остальным окрасам локуса агути. Однако до сих пор неясно, принадлежит ли ген рецессивного черного окраса к этому же локусу или нет.
Ильин (1941) скрещивал волка с черной собакой и получил 7 серых и 6 черных отпрысков. При скрещивании серых потомков между собой было получено 17 серых и 3 черноподпалых, а при скрещивании черных — 12 черных и 3 черноподпалых. Чтобы объяснить полученные данные Ильин предположил существование аллеля черного окраса a
l, который рецессивен но отношению к А, но доминантен к a
t. Тогда волк, в данном случае, должен иметь генотип Aa
l, а черная собака a
la
t. В таком случае, сразу же две случайно отобранные особи имеют очень редкий аллель, что представляется весьма маловероятным. Однако, эти же результаты можно объяснить, если допустить, что волк имел генотип Aa
t, а собака — A
sa
t.
Аллели протяженности окраса
Предназначение этой серии аллелей может показаться странным, однако оно базируется на том, что эти аллели, как и А — аллели связаны с распределением черной и рыжей пигментации. Возможно, что эта серия аллелей контролирует наличие, отсутствие и протяженность только черной пигментации по поверхности туловища.
К настоящему времени известно три аллеля этого локуса.
Фенотип—
Символ
Тигровость — E
br
Нормальное распространение черного пигмента, обусловленное действием других локусов — E
Полное отсутствие черной пигментации шерсти — e
Исходный ген дикого типа Е отвечает за распространение или продуцирование черного пигмента по поверхности всего тела, например, у чисто черных, чепрачных или черноподпалых собак. Гены Е и е являются мутантными аллелями гена Е. Ген E
brотвечает за чередование черных и рыжих полос, что характерно для многих пород. Такой окрас называется тигровым. Ген E
brдоминантен по отношению к гену Е. Ген е отвечает за образование только желтого пигмента по всей поверхности тела и препятствует синтезу черного пигмента в волосе, не влияя на черную пигментацию мочки носа, кожи, губ, пасти и век.
Литтл (1957) опубликовал убедительные доказательства того, что ген тигровости (E
br) доминирует над геном Е, а не наоборот, как считалось раньше. Поэтому, символ e
brдля гена тигровости можно считать устаревшим.
Взаимодействие этих трех аллелей с точки зрения доминантности пока окончательно не выяснено. Условно можно допустить, что каждый аллель полностью доминантен по отношению к нижеследующему в представленной таблице (см. выше). Многочисленные вариации экспрессии гена тигровости позволяют предположить о неполном доминировании гена E
br. Тигровость может варьировать от почти черного окраса, с почти незаметными рыжими пестринами, до песочного, слегка украшенного черными штрихами. С одной стороны, большинство этих вариаций обязано действию модифицирующих полигенов, с другой стороны, усредненный вариант соотношения светлых и темных полос показан для гомозигот E
brE
br. Гетерозиготы E
brE и E
bre имеют отклонения от среднего распределения. Особенно генотип E
bre вызывает уменьшение количества темных полос.
Несмотря на интенсивность пигментации все желтовато-коричневые, желтые, рыжие, красные животные содержат в волосах только желтый пигмент. Реальная степень выраженности окраски зависит от действия модифицирующих генов независимо от того, какой ген присутствует в геноме: А
уили е, или оба вместе.
Литтл (1957) предположил, что наличие маски является доминантным признаком и определяется аллелем из серии Е. Он описал маску как «суперразвитие черного пигмента» обозначив соответствующий ген как E
m. Однако, в литературе нет никаких очевидных подтверждений его предположения. В настоящее время благоразумнее относиться с опаской к такому утверждению. Наследуемость маски мы продолжим обсуждать ниже.
Взаимодействие локусов А и Е
Важность этих двух серий аллелей будет показана в данном разделе. Обе серии контролируют распределение черного и желтого пигментов по шерсти у большинства пород собак. Во взаимодействии эти локусы образуют множество хорошо известных окрасов.
Экспрессия аллелей агути зависит от присутствия гена Е, определяющего насколько полно распространяется черный пигмент по шерстному покрову животного под действием какого-либо из генов агути. Такие генотипы могут быть описаны, как:
Окрас—
Генотип
Сплошной черный — A
s-Е-
A
y-рыжий — A
y-Е-
Чепрачный — a
sa-Е-
Подпалый — a
ta
t-Е-
При мутации гена Е к е, весь пигмент волос становится желтым, т.е. прекращается синтез черного пигмента. При этом невозможно становится «выявить экспрессию агутиаллелей, т к. их экспрессия зависит от присутствия черного пигмента. Следовательно, все вышеперечисленные генотипы, при замене E на е, будут давать только рыжий окрас различной степени интенсивности.
Окрас — Генотип
e-рыжий — A
s-ee
— A
y-ee
— A
sa-ee
— a
ta
tee
Описывая ранее каждый из сравниваемых локусов, мы выяснили, что у собак существует две независимые системы, отвечающие за рыжий окрас. Обе эти системы ведут себя рецессивно по отношению к черному окрасу. Так, А
у-рыжий окрас получается при скрещивании двух черных особей с генотипами A
sA
yEE; а е-рыжий — при скрещивании собак любого окраса из агути-серии, но гетерозиготных по локусу Е (Ее). Со сто процентной уверенностью, A
y-рыжий можно отличить от е-рыжего, когда от двух рыжих родителей получаются еще и чепрачный или черно-подпалый окрасы (a
sa-ЕЕ или a
ta
tEE соотв.), как это часто бывает у такс.
У многих особей, окрашенных по типу А
у, обнаруживается варьирующее количество черного пигмента в шерсти, обычно в виде темно-типированных волос на голове, вдоль спины, на плечах и боках. Если такие волосы многочисленны, то получается соболиный окрас. Для А
у-рыжего окраса характерно также наличие черной маски, захватывающей морду и уши. Напротив, e-рыжий окрас не имеет заметных типированных волос или маски. Однако, некоторые А
у-рыжие особи бывают настолько лишены черных волос, что фенотипически неотличимы от e-рыжего окраса.
Если в одной породе встречаются случаи, когда от двух рыжих особей получаются черные щенки, можно утверждать, что генофонд данной породы имеет как А
у, так и е. Например, при скрещивании рыжей собаки с генотипом A
yA
yEE и рыжей собаки с генотипом A
sA
see потомство будет черным (генотип A
sA
yEe), так как A
sи E доминантны над A
yи е соответственно. А
у-рыжий не может иметь ген A
s, следовательно, этот аллель привнесен от е-рыжей особи. Т.о. это доказывает, что скрещиваемые рыжие собаки имеют разное генетическое происхождение своего окраса.
Было бы интересно узнать соотношение описанных двух типов рыжего окраса в одной породе. Однако, на данный момент времени нет никаких систематических данных. Тем не менее, можно высказать некоторые предположения. Скорее всего большинство рыжих особей несут ген А
у. Фактически, любой рыжий окрас, который имеет некоторое количество черных волос на ушах или вдоль спины, скорее образован за счет действия гена А
у, чем е.
Более четко выявить ген е можно: 1) при скрещивании чепрачного или черно-подпалого окрасов друг с другом в разных сочетаниях, если их генотипы a
sa-Ее или a
ta
tEe. Тогда, генотип всех рыжих потомков первого поколения будет ее; 2) при скрещивании рыжих с чепрачными или черно-подпалыми: если получаются черные щенки, то рыжий производитель должен быть A
s-ее; 3) при скрещивании двух рыжих собак. Как было описано, выше, если получаются черные щенки (A
sA
yEe), то один из родителей должен быть ее. В данном случае проблема состоит в том, чтобы определить, какой из производителей несет ее.
Литтл (1957) уже обсуждал вопрос о том, как распределяются генетически различные рыжие окрасы по породам. Он предположил, что генотип ее несут следующие породы:
Бигль
Далматин
Английский сеттер
Золотой ретривер
Сеттер гордон
Ирландский сеттер
Лабрадор ретривер
Пойнтер
Пудель
Бернс и Фразер (1966) полагают, что золотистый кокер-спаниель тоже может нести ее, но Литтл (1957) считает, что в этой породе одновременно существуют и А
у-рыжие и ее-рыжие особи. Присутствие обоих генотипов в одной породе может вызвать путаницу у собаководов с начальными генетическими знаниями, которые не представляют, что могут одновременно существовать два генетически различных рыжих окраса. К таким породам Литтл (1957) отнес следующие:
чау-чау
английский спаниель
филд-спанпель
Аллель тигровости, E
brпроявляет свои характерные черты только на рыжих областях, расположение и площадь которых определяется генами серии агути. Ген E
brиндуцирует синтез черного пигмента в волосах, расположенных на рыжих ареалах в виде пигментированных полос или штрихов на рыжем фоне.
Окрас—
Генотип
Сплошной черный — A
s-E
br-
Тигровый — A
y-E
br-
Темно-тигровый — a
sa-E
br-
Черно-тигровый — a
ta
tE
br-
Ген A
sэпистатичен по отношению к E
br. Это не удивительно, т.к. A
s-индивидуумы полностью черные и не имеют рыжих участков, на которых могли бы развиваться тигровины. Однако, в комбинации с геном А
у, A
brэкспрессируется.
Несомненно, что у большинства пород тигровый окрас определяется генотипом A
y-E
br-.
Генотип a
sa-E
br-, определяющий чепрачно-тигровый окрас, дает тигровость на рыжих участках типично чепрачного окраса. Расположение полос будет зависеть от типа чепрачности. Если развитие чепрака незначительно, то полосы проявятся на большей части тела, хотя сами по себе они будут темными. Если чепрак развит значительно и охватывает большую часть туловища, то полосы останутся только на ногах и животе, и такая собака будет выглядеть, как очень темно-тигровая.
Генотип a
ta
tE
br-, определяющий темно-тигровый окрас, дает полосы, расположенные на подпалах. Фенотип таких животных — наиболее четкое доказательство того, что E
brработает только на рыжем фоне. Эти собаки, несомненно, черно-подпалые, несмотря на тигровость подпалов, интенсивность которой может варьировать.
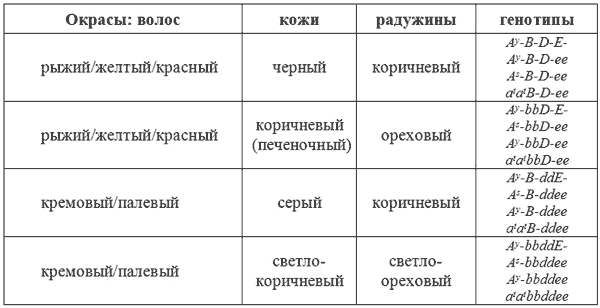
Коричневый окрас
Организм млекопитающих продуцирует всего два пигмента — это желтый и черный пигменты. Они содержатся в гранулах, расположенных внутри волоса. Цвет, количество и форма этих гранул и определяют окрас волоса. Черный волос имеет овальные, интенсивно окрашенные гранулы, содержащие черный пигмент меланин. Палевые, рыжие, красные или желтые волосы имеют более мелкие, округлые гранулы, содержащие желтый пигмент.
У животных коричневого окраса (иногда его называют шоколадный или печеночный) или коричнево-чепрачных, или коричнево-подпалых, гранулы в коричневых волосах имеют меньше черного пигмента, чем в черных волосах, и оптически выглядят как коричневые. У тех же животных рыжие волосы не изменены. Очевидно, что мутантный ген, продуцирующий коричневый цвет, действует только на гранулы, содержащие черный пигмент, осветляя их. Для человеческого глаза это выглядит как коричневый. Т.о, здесь мы имеем дело с двумя альтернативными генами, определяющими черный окрас волос (В) и коричневый окрас волос (b). Исходным является ген B. Он доминантен по отношению к мутантному гену b. Ген b изменяет все черные волосы на коричневые, а так же осветляет все пигментированные участки кожи губ, пасти, когтей, мочки носа и радужину глаз.
Сплошной коричневый окрас получается при комбинации гена b с геном A
sиз агути-серки (A
s-bb). В следующем разделе, касающемся гена ослабления пигментации, вариации коричневого окраса будут обсуждаться более полно.
Т.к. ген b не влияет на пигментацию рыжих районов, то можно всех рыжих собак разделить на два типа: это «черные» рыжие собаки, несущие ген В в генотипе А
у-В- или B-ее, и «коричневые» рыжие собаки, несущие b: A
y-bb или bbee. Эти окрасы внешне не идентичны, так как во втором случае происходит ослабление пигментации кожи и глаз и такие собаки при рыжем окрасе будут иметь коричневую мочку нос, обводку глаз, губ, осветленные глаза и пр.
Вообще, для собак не характерны случаи, когда от коричневых родителей получали черных щенков. Однако, Франклинг (1971) описала случай появления пяти полностью черных пометов при скрещивании семи пар различных коричневых далматинов. При этом она подчеркнула, что владельцы заслуживают полного доверия, а скрещивания происходили под контролем специалистов.
Если это действительно так, то можно предположить, что шоколадный окрас получается не только в результате мутации B в b, но и в результате какой-то неизученной пока мутации другого независимого локуса. Такая ситуация чрезвычайно редка в генетике млекопитающих и известна только у американских норок. Поэтому, приведенные Франклинг сведения не получили всеобщего признания.
Ослабление окраса
Пигментные гранулы в нормально окрашенном волосе расположены достаточно непрерывно по мере роста волос. В основании цвет менее интенсивен. Например, черные волосы имеют интенсивно черный окрас на концах, бледнея к основанию и становясь совсем голубоватыми у корня, благодаря постепенному уменьшению количества гранул по направлению к коже. То же самое происходит и с пигментацией рыжих волос. Они интенсивно пигментированы на концах, бледнея до палевого у основания. При внешнем осмотре более бледные основания полос не видны, но заметны у стриженых или длинношерстных особей.
«Голубизна» голубых собак тоже является результатом уменьшения количества пигментных гранул, однако это обусловлено другими механизмами. Гранулы в волосах таких собак откладываются не непрерывно, а островками или глыбками. Вследствие этого, одни участки волос содержат больше пигмента, другие меньше. В результате этого, для человеческого глаза шерсть, состоящая из таких волос, будет выглядеть серой в случае черного пигмента или палевой в случае желтого пигмента. Различия в диспозиции пигментных гранул обязаны паре генов: D и d. Последний вызывает прерывистое распределение пигментных гранул в волосе. Таким образом, ген D отвечает за интенсивное прокрашивание, а ген d — за ослабление пигментации.
У млекопитающих существует четыре основных окраса, именуемые как черный, голубой, коричневый и лиловый У собак встречаются все четыре окраса и получаются они в результате комбинаций генов b и d с A
sи Е.
Окрас — Генотип
Черный — A
s-B-D-E-
Голубой — A
s-B-ddE-
Коричневый — A
s-bbD-E-
Лиловый — A
s-bbddE-
У многих пород собак встречаются и другие окрасы. Если вместо A
s- в геноме присутствуют a
ta
tте же самые четыре основных окраса дают различных двуцветных или подпалых собак.
Окрас—
Генотип Черно-подпалый — a
ta
tB-D-E-
Голубо-подпалый — a
ta
tB-ddE-
Коричнево-подпалый — a
ta
tbbD-E-
Лилово-подпалый — a
ta
tbbddE-
Ген d влияет как на черную (коричневую), так и на желтую пигментацию. Следовательно, голубо-подпалый и лилово-подпалый корректнее было бы обозначить как голубо-кремовый и лилово-кремовый. Однако, приставку -подпалый следует сохранять в тех случаях, когда нужно подчеркнуть определенный характер распределения цветов.
Ген d присутствует у огромного количества пород собак и распространенность его зависит от степени популярности голубого окраса Ген b не так распространен, но встречается особенно часто у спаниелей. Действительно, некоторые породы спаниелей всегда гомозиготны по гену о (bb). Это американский и ирландский водяной спаниели и Суссекс. У других пород разрешены как ген В, так и b. Любопытно, что породы, имеющие ген b, как правило, не имеют гена d, следовательно лиловая комбинация (bbdd) крайне редко встречается. Однако, этот генотип формирует основу уникального окраса веймаранской гончей, генотип окраса которой: A
s-bbddE-.
Большинство любителей-собаководов имеют свои предпочтительные термины для обозначения различных окрасов. Так, коричневый окрас часто называют печеночным или шоколадным. Лиловый окрас настолько редок и необычен, что для него нет общего термина. Окрас веймаранской гончей описывается как серебристый, что является по общему признанию наиболее подходящим термином. Однако, для собаководов полезно знать, что в генетике млекопитающих термин «лиловый» обозначает окрас, обусловленный генотипом bbdd.
В комбинации с генами А
уи E
brгены В и D и их аллели дают четыре типа тигровых собак.
Окрас—
Генотип Черно-тигровый — A
y-B-D-E
br-
Голубо-тигровый — A
y-B-ddE
br-
Коричнево-тигровый — A
y-bbD-E
br-
Лилово-тигровый — A
y-bbddE
br-
Сказанное применимо ко всем формам тигровости, описанным ранее. Сомнительно, чтобы некоторые коричневые и лиловые формы отчетливо распознавались собаководами. В любом случае они были бы необычны.
Ген b действует только на черный пигмент, следовательно, все рыжие фенотипы выглядят обычно, но присутствие этого гена выявляется по изменениям окраски радужины глаз, кожи, когтей. Ген d ослабляет рыжий до кремового, при этом изменения пигментации кожи, глаз и когтей незначительны.
Соболиный окрас имеет генотип A
y-B-D-E-. Интенсивность затенения зависит от наличия модифицирующих полигенов. Причем, черное затенение может быть ослаблено до коричневого под воздействием гена b, тогда получится шоколадно-соболиный окрас. Однако, сомнительно, чтобы такой окрас можно было легко распознать. Многие собаководы определяют такой окрас как желтый или рыжий, особенно, если затенение незначительно и тогда оно не выделяется на рыжем фоне. Соответственно, голубо-соболиный или лилово-соболиный сразу и не отличить от простого кремового или палевого.
Аллели белого окраса, альбинизма
Серия аллелей белого окраса или альбинотическая фундаментально контролирует синтез пигмента по всему телу. Можно только удивляться, почему целая серия генов так названа, хотя альбинизм чрезвычайно редок среди собак. Ответ следует искать в общей генетике окрасов млекопитающих. Альбинизм может быть редок у собак, но он обычен у многих млекопитающих. Локус альбинизма более мутабилен, чем другие локусы. Таким образом, появилась серия аллелей с характерными фенотипами. Более того, эти фенотипы сходны у всех видов млекопитающих, поэтому общие знания хорошо применимы и в случаях собак, так как явление альбинизма у них крайне редко.
Общее количество мутантных аллелей локуса альбинизма неизвестно, но, по крайней мере, три из них установлены.
Фенотипическое проявление - Символы гена
Полное проявление окраса — С
Шиншилла — c
ch
Голубоглазый альбинизм — c
b
Полный альбинизм — c
Все нормально окрашенные собаки имеют ген С, который разрешает экспрессию генов синтеза пигментов. Типично, что аллели, следующие ниже в таблице, разрешают экспрессию все меньшего количества пигмента. Последний ген в таблице полностью запрещает синтез любого пигмента.
Аллель шиншилла (c
ch) характерен тем, что осветляет рыжий до бледно-желтого, и почти или совсем не влияет на развитие черного пигмента. Похоже, что этот ген присутствует у пород, имеющих бледно-желтый окрас, таких, как норвежский элькхунд. Ген c
chосветляет как А
у- рыжих; так и ee-рыжих собак. В частности, наиболее ожидаемым генотипом для золотого ретривера следует признать c
chc
chee. Окрас таких шиншилловых животных может варьировать от теплого кремового до почти белого. Большинство этих вариаций образуются под действием модифицирующих руфус-полигенов. Более подробно о них будет сказано ниже. Литтл же предположил, что значительные вариации этого окраса обязаны существованию более чем одного аллеля шиншилла у собак.
Второй аллель шиншиллы, обозначенный Литтлем, как c
e, возможно, отвечает за почти белый фенотип с кремовой тенью вдоль спины, на плечах и голове. Если принять существования этого аллеля, то генотип этих окрасов будет А
у-c
ec
eили c
ec
eee. Можно также предположить, что белые вариации окраса у некоторых пород имеют эти же генотипы, например, вестхайленд-уайт-терьер.
Ни один из полностью белых окрасов у собак не является настоящим альбиносным, так как при этом собаки имеют темные глаза. Следует заметить, что возможность существования аллеля типа шиншилла c
chосновывается более на фенотипических наблюдениях, чем на результатах экспериментальных скрещиваний.
Существование нескольких аллелей шиншиллы можно подвергнуть сомнению, предположив, что не шиншилла, а модифицирующие полигены ослабляют рыжий до палевого во многих случаях. В конечном итоге, пока следует признать достоверным существование только одного аллеля гена шиншилла.
Литтл часто упоминал в своей книге, что ген С не полностью доминантен по отношению к c
ch. Но это противоречит наблюдениям у других видов животных. Однако, если признать, что в случаях, описанных Литтлем, рыжий окрас ослабляется под действием полигенов, а не c
ch, тогда неполное доминирование С не удивительно.
Персон и Ашер (1929) описали два очень светло-окрашенных фенотипа со светло-голубыми глазами и тускло-красными зрачками. Ряд исследователей считают, что, по крайней мере, один из них несет аллель альбинизма, который был назван как «голубоглазый альбинизм» (c
b). О наследуемости c
bочень мало известно. Предложено поместить этот ген между c
chи с в представленной выше таблице.
Полный альбинизм, выраженный в чисто белой шерсти, розовых зрачках и просвечивающей красной радужине, крайне редок у собак. Витни (1947) и Литтл (1957) описали таких альбиносов у пекинесов.
Ослабление окраса, сопряженное с розовоглазием
Эта форма ослабления, в целом, обычна для млекопитающих, но редка у собак. Возможно, что одна из форм осветленного фенотипа с голубыми глазами, описанная Персоком и Ашером (см. выше) имела этот ген. Для подобного типа ослабления характерны голубоватый или сероватый окрас и красноватые глаза. Его следует отличать от обычного ослабления до голубого (ген d), когда зрачки остаются темными. Несмотря на уменьшение количества пигмента, «розовоглазое ослабление» не связано с локусом альбинизма, а является мутантной формой (р) гена Р, также отвечающего за развитие интенсивной окраски шерсти и глаз.