Прежде, чем приступить к изложению генетики пород собак, необходимо в общих чертах ознакомиться с тем, что известно о генах, определяющих окрасы и структуру шерсти у собак, и их действии. Для этого было бы полезно иметь представление об общих принципах наследственности, с которыми можно ознакомиться в популярных, и специальных изданиях по генетике. В настоящей главе мы обсудим действие генов более детально и систематизировано. В основном использовались общепринятые в генетике млекопитающих символы для обозначения генов и их мутантов.
До сих пор было проведено ничтожно мало контролируемых экспериментов по скрещиванию собак. Такая работа требует значительных расходов и длительного времени. Поэтому, в некоторых случаях использованы результаты случайных наблюдений. Однако, основная масса информации по генетике собак была получена из достоверных источников.
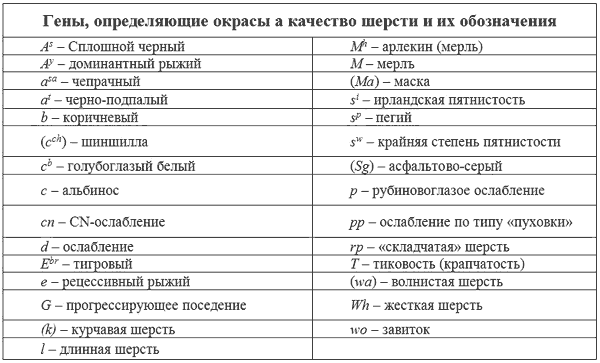
Аллели «Агути»
Ряд наиболее распространенных фенотипов окраса определяются серией аллелей, известной как «агути». Этот термин произошел от названия небольшого грызуна с коричневосерой шерстью, которая прекрасно скрывает его от хищников. У псовых соответствующий окрас называют волче-серым (зонарным). Однако, в этом случае, такой окрас позволяет хищнику незаметно подкрадываться к жертве.
В генетике собак термин «агути» позаимствован из генетики грызунов, где ген «агути» хорошо изучен. Выяснено, что исходный (дикий тип) ген «агути» является родоначальником целой серии аллелей. В настоящее время выявлены следующие аллели:
Фенотипическое проявление—
Символ
Сплошной черный — A
s
Доминантный желтый (рыжий) — A
y
Агути (волче-серый) — A
Чепрачный — a
sa
Черно-подпалый — a
t
Исходный ген А («агути») отвечает за волче-серый окрас диких псовых и большинства, если не всех пород собак, имеющих волчеобразный серый окрас. К таким породам можно отнести серую сибирскую хаски, серого норвежского элькхунда и других собак крайнего Севера. Возможно, что серая немецкая овчарка имеет генотип АА.
В результате мутагенеза ген А дал четыре мутантных аллеля: два доминантных (A
sи A
y) и два рецессивных (a
saи a
t) по отношению к исходному гену А.
Ген A
sотвечает за сплошной черный окрас, характерный для многих пород. Этот окрас варьирует от угольно-черного до буро-чёрного. Последний образуется в результате различного числа остаточных агути-подобных волос. Нечистокровные черные собаки часто имеют бурый оттенок. При ближайшем рассмотрении их шерсти можно выявить некоторое количество зонарных волос наряду с чисто черными. Наиболее простое объяснение этому явлению заключается с том, что ген A
sсам по себе не может индуцировать образование чисто черных волос. Для этого ему требуется наличие достаточного количества модифицирующих полигенов, селекционно отобранных при выведении чистокровных пород. Можно также предположить, что ген A
sне полностью доминантен, тогда генотип A
sA
sопределяет чисто черный окрас, а генотипы A
sA
yили A
sA — бурый. Агути-подобные волосы могут давать также специфический красноватый оттенок, описанный Литтлем (1957), который предположил, что такой отгонок образуется благодаря неполному доминированию A
sнад A
yи a
t. Литтл показал, что у особей A
sA
yкрасноватый оттенок встречается в основном на боках, голове, шее и конечностях, а особи с генотипом A
sa
tимеют красноватый оттенок только там, где должны располагаться подпалы у особей a
ta
t. Такое неполное доминирование вполне возможно. Однако, следует заметить, что статистически это еще не доказано.
Доминантный аллельный ген A
yотвечает за рыжий окрас собаки. Этот ген дает чисто рыжих собак, но при этом обнаруживаются волосы с черными кончиками (т.н. типированные) на голове, плечах, вдоль спины, включая хвост. При наличии значительного количества таких волос можно говорить о соболином окрасе. Поэтому, этот аллель можно назвать также аллелью соболиного или соболе-рыжего окраса. По этой терминологии полностью, рыжие формы можно обозначить как золотисто-соболиные или светло-соболиные. Количество зачерненных волос или степень проявления соболиного окраса варьирует, что обусловлено полигенами, которые наследуются независимо от А. В совокупности, эти полигены известны под названием «затеняющие».
Литтл (1957) предположил, что соболиный окрас проявляется при гетерозиготном генотипе A
ya
t. Тогда, при скрещивании обоих особей соболиного окраса, потомство должно быть рыжим, соболиным и черно-подпалым в соотношении 1:2:1 соответственно. Экспериментально, примерно так и получалось. Однако, и от чисто рыжих собак также получались периодически черно-подпалые щенки, что говорит о гетерозиготности обоих родителей (A
ya
t). Это обстоятельство в значительной степени опровергает идею неполного доминирования A
yнад a
t. Можно предположить, что полигены затенения сами по себе так взаимодействуют с генами a
tи A
y, что в случае гетерозиготы A
ya
tполучается темно-соболиной окрас, а в случае гомозиготы A
yA
y— рыжий окрас со слабым или незначительным затенением. Но это весьма сомнительно.
Ген чепрачности a
saопределяет V-образный характер пигментации шерсти по обеим сторонам туловища, как, например, у эрдельтерьера или бигля. На первый взгляд, фенотипы a
saи a
tсходны, но чепрачные особи имеют больше рыжего, чем черно-подпалые, особенно на морде, плечах, боках и конечностях. Однако, большое распространение черного в чепрачном окрасе может напоминать черно-подпалый. Из двух этих окрасов чепрачный наиболее вариабелен.
В типичном случае черно-подпалого окраса (доберманы, например) черный распространяется на всю верхнюю часть туловища, а рыжий ограничен внутренней поверхностью конечностей, грудными отметинами и нижней поверхностью морды. Два характерных пятна расположены над глазами. Для чепрачного окраса характерны возрастные изменения. Так, чепрачные щенки рождаются очень похожими на черноподпалых, с возрастом они светлеют, рыжие отметины увеличиваются по площади, пока молодые животные не становятся истинно чепрачными. Нельзя не отметить интригующий факт, что соболиные собаки подвержены сходным возрастным изменениям. Это явление настолько всеобщее, что и детеныши волка имеют более темный окрас, чем взрослые животные.
Интересно, что у большинства чистокровных пород собак, за исключением эскимосских и скандинавских, аллель дикого типа А отсутствует. Такое отсутствие столь характерно, что кажется преднамеренным. Возможно, что древние селекционеры стремились закреплять такие цветовые вариации, которые, во-первых, отличали бы домашних собак от диких форм и, во-вторых, служили бы символом одомашнивания собак.
Наиболее очевидно, что самым близким аллелем к гену А является аллель a
sa. В основном А и a
saпроявляются сходно. Так, у нечистокровных животных наблюдаются все переходы от зонарного (волче-серого) до чепрачного окрасов. Более того, Фокс (1978) предположил, что зонарный и чепрачный окрасы обусловлены действием одного и того же гена. Он скрещивал койота и бигля и в последующих двух поколениях получил в общей сложности 16 животных, которые летом были темно-соболиного окраса, а зимой светлели до светло-соболиного. Однако, по рисункам, представленным Фоксом, потомство было скорее зонарным или светло-чепрачным. В любом случае, это наблюдение показательно, но не убедительно.
Литтл (1957) предположил, что чепрачный и черно-подпалый окрасы обусловлены действием одного аллеля, а различия между ними являются результатом работы модифицирующих полигенов. Однако, в 1976 году Виллис убедительно продемонстрировал различия между генами a
saи a
tи доказал доминирование a
saнад a
t. Не очень правдоподобна также и идея о существовании более одного аллеля, кодирующего чепрачный окрас. В этом случае наблюдались бы явные скачкообразные различия, тогда как встречаются постепенные переходы от самых затемненных до самых светлых чепрачных окрасов.
Кажется возможным, что у немецкой овчарки черный окрас кодируется не геном A
s. Так, Виллис описал черную форму, рецессивную по отношению ко всем остальным окрасам локуса агути. Однако до сих пор неясно, принадлежит ли ген рецессивного черного окраса к этому же локусу или нет.
Ильин (1941) скрещивал волка с черной собакой и получил 7 серых и 6 черных отпрысков. При скрещивании серых потомков между собой было получено 17 серых и 3 черноподпалых, а при скрещивании черных — 12 черных и 3 черноподпалых. Чтобы объяснить полученные данные Ильин предположил существование аллеля черного окраса a
l, который рецессивен но отношению к А, но доминантен к a
t. Тогда волк, в данном случае, должен иметь генотип Aa
l, а черная собака a
la
t. В таком случае, сразу же две случайно отобранные особи имеют очень редкий аллель, что представляется весьма маловероятным. Однако, эти же результаты можно объяснить, если допустить, что волк имел генотип Aa
t, а собака — A
sa
t.
Аллели протяженности окраса
Предназначение этой серии аллелей может показаться странным, однако оно базируется на том, что эти аллели, как и А — аллели связаны с распределением черной и рыжей пигментации. Возможно, что эта серия аллелей контролирует наличие, отсутствие и протяженность только черной пигментации по поверхности туловища.
К настоящему времени известно три аллеля этого локуса.
Фенотип—
Символ
Тигровость — E
br
Нормальное распространение черного пигмента, обусловленное действием других локусов — E
Полное отсутствие черной пигментации шерсти — e
Исходный ген дикого типа Е отвечает за распространение или продуцирование черного пигмента по поверхности всего тела, например, у чисто черных, чепрачных или черноподпалых собак. Гены Е и е являются мутантными аллелями гена Е. Ген E
brотвечает за чередование черных и рыжих полос, что характерно для многих пород. Такой окрас называется тигровым. Ген E
brдоминантен по отношению к гену Е. Ген е отвечает за образование только желтого пигмента по всей поверхности тела и препятствует синтезу черного пигмента в волосе, не влияя на черную пигментацию мочки носа, кожи, губ, пасти и век.
Литтл (1957) опубликовал убедительные доказательства того, что ген тигровости (E
br) доминирует над геном Е, а не наоборот, как считалось раньше. Поэтому, символ e
brдля гена тигровости можно считать устаревшим.
Взаимодействие этих трех аллелей с точки зрения доминантности пока окончательно не выяснено. Условно можно допустить, что каждый аллель полностью доминантен по отношению к нижеследующему в представленной таблице (см. выше). Многочисленные вариации экспрессии гена тигровости позволяют предположить о неполном доминировании гена E
br. Тигровость может варьировать от почти черного окраса, с почти незаметными рыжими пестринами, до песочного, слегка украшенного черными штрихами. С одной стороны, большинство этих вариаций обязано действию модифицирующих полигенов, с другой стороны, усредненный вариант соотношения светлых и темных полос показан для гомозигот E
brE
br. Гетерозиготы E
brE и E
bre имеют отклонения от среднего распределения. Особенно генотип E
bre вызывает уменьшение количества темных полос.
Несмотря на интенсивность пигментации все желтовато-коричневые, желтые, рыжие, красные животные содержат в волосах только желтый пигмент. Реальная степень выраженности окраски зависит от действия модифицирующих генов независимо от того, какой ген присутствует в геноме: А
уили е, или оба вместе.
Литтл (1957) предположил, что наличие маски является доминантным признаком и определяется аллелем из серии Е. Он описал маску как «суперразвитие черного пигмента» обозначив соответствующий ген как E
m. Однако, в литературе нет никаких очевидных подтверждений его предположения. В настоящее время благоразумнее относиться с опаской к такому утверждению. Наследуемость маски мы продолжим обсуждать ниже.
Взаимодействие локусов А и Е
Важность этих двух серий аллелей будет показана в данном разделе. Обе серии контролируют распределение черного и желтого пигментов по шерсти у большинства пород собак. Во взаимодействии эти локусы образуют множество хорошо известных окрасов.
Экспрессия аллелей агути зависит от присутствия гена Е, определяющего насколько полно распространяется черный пигмент по шерстному покрову животного под действием какого-либо из генов агути. Такие генотипы могут быть описаны, как:
Окрас—
Генотип
Сплошной черный — A
s-Е-
A
y-рыжий — A
y-Е-
Чепрачный — a
sa-Е-
Подпалый — a
ta
t-Е-
При мутации гена Е к е, весь пигмент волос становится желтым, т.е. прекращается синтез черного пигмента. При этом невозможно становится «выявить экспрессию агутиаллелей, т к. их экспрессия зависит от присутствия черного пигмента. Следовательно, все вышеперечисленные генотипы, при замене E на е, будут давать только рыжий окрас различной степени интенсивности.
Окрас — Генотип
e-рыжий — A
s-ee
— A
y-ee
— A
sa-ee
— a
ta
tee
Описывая ранее каждый из сравниваемых локусов, мы выяснили, что у собак существует две независимые системы, отвечающие за рыжий окрас. Обе эти системы ведут себя рецессивно по отношению к черному окрасу. Так, А
у-рыжий окрас получается при скрещивании двух черных особей с генотипами A
sA
yEE; а е-рыжий — при скрещивании собак любого окраса из агути-серии, но гетерозиготных по локусу Е (Ее). Со сто процентной уверенностью, A
y-рыжий можно отличить от е-рыжего, когда от двух рыжих родителей получаются еще и чепрачный или черно-подпалый окрасы (a
sa-ЕЕ или a
ta
tEE соотв.), как это часто бывает у такс.
У многих особей, окрашенных по типу А
у, обнаруживается варьирующее количество черного пигмента в шерсти, обычно в виде темно-типированных волос на голове, вдоль спины, на плечах и боках. Если такие волосы многочисленны, то получается соболиный окрас. Для А
у-рыжего окраса характерно также наличие черной маски, захватывающей морду и уши. Напротив, e-рыжий окрас не имеет заметных типированных волос или маски. Однако, некоторые А
у-рыжие особи бывают настолько лишены черных волос, что фенотипически неотличимы от e-рыжего окраса.
Если в одной породе встречаются случаи, когда от двух рыжих особей получаются черные щенки, можно утверждать, что генофонд данной породы имеет как А
у, так и е. Например, при скрещивании рыжей собаки с генотипом A
yA
yEE и рыжей собаки с генотипом A
sA
see потомство будет черным (генотип A
sA
yEe), так как A
sи E доминантны над A
yи е соответственно. А
у-рыжий не может иметь ген A
s, следовательно, этот аллель привнесен от е-рыжей особи. Т.о. это доказывает, что скрещиваемые рыжие собаки имеют разное генетическое происхождение своего окраса.
Было бы интересно узнать соотношение описанных двух типов рыжего окраса в одной породе. Однако, на данный момент времени нет никаких систематических данных. Тем не менее, можно высказать некоторые предположения. Скорее всего большинство рыжих особей несут ген А
у. Фактически, любой рыжий окрас, который имеет некоторое количество черных волос на ушах или вдоль спины, скорее образован за счет действия гена А
у, чем е.
Более четко выявить ген е можно: 1) при скрещивании чепрачного или черно-подпалого окрасов друг с другом в разных сочетаниях, если их генотипы a
sa-Ее или a
ta
tEe. Тогда, генотип всех рыжих потомков первого поколения будет ее; 2) при скрещивании рыжих с чепрачными или черно-подпалыми: если получаются черные щенки, то рыжий производитель должен быть A
s-ее; 3) при скрещивании двух рыжих собак. Как было описано, выше, если получаются черные щенки (A
sA
yEe), то один из родителей должен быть ее. В данном случае проблема состоит в том, чтобы определить, какой из производителей несет ее.
Литтл (1957) уже обсуждал вопрос о том, как распределяются генетически различные рыжие окрасы по породам. Он предположил, что генотип ее несут следующие породы:
Бигль
Далматин
Английский сеттер
Золотой ретривер
Сеттер гордон
Ирландский сеттер
Лабрадор ретривер
Пойнтер
Пудель
Бернс и Фразер (1966) полагают, что золотистый кокер-спаниель тоже может нести ее, но Литтл (1957) считает, что в этой породе одновременно существуют и А
у-рыжие и ее-рыжие особи. Присутствие обоих генотипов в одной породе может вызвать путаницу у собаководов с начальными генетическими знаниями, которые не представляют, что могут одновременно существовать два генетически различных рыжих окраса. К таким породам Литтл (1957) отнес следующие:
чау-чау
английский спаниель
филд-спанпель
Аллель тигровости, E
brпроявляет свои характерные черты только на рыжих областях, расположение и площадь которых определяется генами серии агути. Ген E
brиндуцирует синтез черного пигмента в волосах, расположенных на рыжих ареалах в виде пигментированных полос или штрихов на рыжем фоне.
Окрас—
Генотип
Сплошной черный — A
s-E
br-
Тигровый — A
y-E
br-
Темно-тигровый — a
sa-E
br-
Черно-тигровый — a
ta
tE
br-
Ген A
sэпистатичен по отношению к E
br. Это не удивительно, т.к. A
s-индивидуумы полностью черные и не имеют рыжих участков, на которых могли бы развиваться тигровины. Однако, в комбинации с геном А
у, A
brэкспрессируется.
Несомненно, что у большинства пород тигровый окрас определяется генотипом A
y-E
br-.
Генотип a
sa-E
br-, определяющий чепрачно-тигровый окрас, дает тигровость на рыжих участках типично чепрачного окраса. Расположение полос будет зависеть от типа чепрачности. Если развитие чепрака незначительно, то полосы проявятся на большей части тела, хотя сами по себе они будут темными. Если чепрак развит значительно и охватывает большую часть туловища, то полосы останутся только на ногах и животе, и такая собака будет выглядеть, как очень темно-тигровая.
Генотип a
ta
tE
br-, определяющий темно-тигровый окрас, дает полосы, расположенные на подпалах. Фенотип таких животных — наиболее четкое доказательство того, что E
brработает только на рыжем фоне. Эти собаки, несомненно, черно-подпалые, несмотря на тигровость подпалов, интенсивность которой может варьировать.
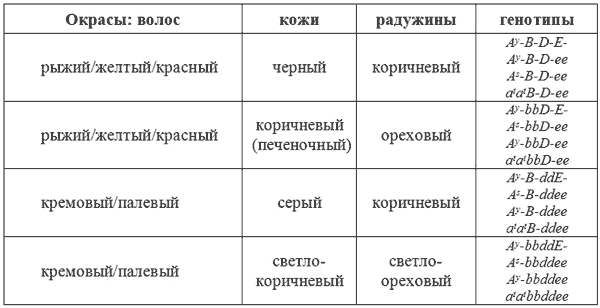
Коричневый окрас
Организм млекопитающих продуцирует всего два пигмента — это желтый и черный пигменты. Они содержатся в гранулах, расположенных внутри волоса. Цвет, количество и форма этих гранул и определяют окрас волоса. Черный волос имеет овальные, интенсивно окрашенные гранулы, содержащие черный пигмент меланин. Палевые, рыжие, красные или желтые волосы имеют более мелкие, округлые гранулы, содержащие желтый пигмент.
У животных коричневого окраса (иногда его называют шоколадный или печеночный) или коричнево-чепрачных, или коричнево-подпалых, гранулы в коричневых волосах имеют меньше черного пигмента, чем в черных волосах, и оптически выглядят как коричневые. У тех же животных рыжие волосы не изменены. Очевидно, что мутантный ген, продуцирующий коричневый цвет, действует только на гранулы, содержащие черный пигмент, осветляя их. Для человеческого глаза это выглядит как коричневый. Т.о, здесь мы имеем дело с двумя альтернативными генами, определяющими черный окрас волос (В) и коричневый окрас волос (b). Исходным является ген B. Он доминантен по отношению к мутантному гену b. Ген b изменяет все черные волосы на коричневые, а так же осветляет все пигментированные участки кожи губ, пасти, когтей, мочки носа и радужину глаз.
Сплошной коричневый окрас получается при комбинации гена b с геном A
sиз агути-серки (A
s-bb). В следующем разделе, касающемся гена ослабления пигментации, вариации коричневого окраса будут обсуждаться более полно.
Т.к. ген b не влияет на пигментацию рыжих районов, то можно всех рыжих собак разделить на два типа: это «черные» рыжие собаки, несущие ген В в генотипе А
у-В- или B-ее, и «коричневые» рыжие собаки, несущие b: A
y-bb или bbee. Эти окрасы внешне не идентичны, так как во втором случае происходит ослабление пигментации кожи и глаз и такие собаки при рыжем окрасе будут иметь коричневую мочку нос, обводку глаз, губ, осветленные глаза и пр.
Вообще, для собак не характерны случаи, когда от коричневых родителей получали черных щенков. Однако, Франклинг (1971) описала случай появления пяти полностью черных пометов при скрещивании семи пар различных коричневых далматинов. При этом она подчеркнула, что владельцы заслуживают полного доверия, а скрещивания происходили под контролем специалистов.
Если это действительно так, то можно предположить, что шоколадный окрас получается не только в результате мутации B в b, но и в результате какой-то неизученной пока мутации другого независимого локуса. Такая ситуация чрезвычайно редка в генетике млекопитающих и известна только у американских норок. Поэтому, приведенные Франклинг сведения не получили всеобщего признания.
Ослабление окраса
Пигментные гранулы в нормально окрашенном волосе расположены достаточно непрерывно по мере роста волос. В основании цвет менее интенсивен. Например, черные волосы имеют интенсивно черный окрас на концах, бледнея к основанию и становясь совсем голубоватыми у корня, благодаря постепенному уменьшению количества гранул по направлению к коже. То же самое происходит и с пигментацией рыжих волос. Они интенсивно пигментированы на концах, бледнея до палевого у основания. При внешнем осмотре более бледные основания полос не видны, но заметны у стриженых или длинношерстных особей.
«Голубизна» голубых собак тоже является результатом уменьшения количества пигментных гранул, однако это обусловлено другими механизмами. Гранулы в волосах таких собак откладываются не непрерывно, а островками или глыбками. Вследствие этого, одни участки волос содержат больше пигмента, другие меньше. В результате этого, для человеческого глаза шерсть, состоящая из таких волос, будет выглядеть серой в случае черного пигмента или палевой в случае желтого пигмента. Различия в диспозиции пигментных гранул обязаны паре генов: D и d. Последний вызывает прерывистое распределение пигментных гранул в волосе. Таким образом, ген D отвечает за интенсивное прокрашивание, а ген d — за ослабление пигментации.
У млекопитающих существует четыре основных окраса, именуемые как черный, голубой, коричневый и лиловый У собак встречаются все четыре окраса и получаются они в результате комбинаций генов b и d с A
sи Е.
Окрас — Генотип
Черный — A
s-B-D-E-
Голубой — A
s-B-ddE-
Коричневый — A
s-bbD-E-
Лиловый — A
s-bbddE-
У многих пород собак встречаются и другие окрасы. Если вместо A
s- в геноме присутствуют a
ta
tте же самые четыре основных окраса дают различных двуцветных или подпалых собак.
Окрас—
Генотип Черно-подпалый — a
ta
tB-D-E-
Голубо-подпалый — a
ta
tB-ddE-
Коричнево-подпалый — a
ta
tbbD-E-
Лилово-подпалый — a
ta
tbbddE-
Ген d влияет как на черную (коричневую), так и на желтую пигментацию. Следовательно, голубо-подпалый и лилово-подпалый корректнее было бы обозначить как голубо-кремовый и лилово-кремовый. Однако, приставку -подпалый следует сохранять в тех случаях, когда нужно подчеркнуть определенный характер распределения цветов.
Ген d присутствует у огромного количества пород собак и распространенность его зависит от степени популярности голубого окраса Ген b не так распространен, но встречается особенно часто у спаниелей. Действительно, некоторые породы спаниелей всегда гомозиготны по гену о (bb). Это американский и ирландский водяной спаниели и Суссекс. У других пород разрешены как ген В, так и b. Любопытно, что породы, имеющие ген b, как правило, не имеют гена d, следовательно лиловая комбинация (bbdd) крайне редко встречается. Однако, этот генотип формирует основу уникального окраса веймаранской гончей, генотип окраса которой: A
s-bbddE-.
Большинство любителей-собаководов имеют свои предпочтительные термины для обозначения различных окрасов. Так, коричневый окрас часто называют печеночным или шоколадным. Лиловый окрас настолько редок и необычен, что для него нет общего термина. Окрас веймаранской гончей описывается как серебристый, что является по общему признанию наиболее подходящим термином. Однако, для собаководов полезно знать, что в генетике млекопитающих термин «лиловый» обозначает окрас, обусловленный генотипом bbdd.
В комбинации с генами А
уи E
brгены В и D и их аллели дают четыре типа тигровых собак.
Окрас—
Генотип Черно-тигровый — A
y-B-D-E
br-
Голубо-тигровый — A
y-B-ddE
br-
Коричнево-тигровый — A
y-bbD-E
br-
Лилово-тигровый — A
y-bbddE
br-
Сказанное применимо ко всем формам тигровости, описанным ранее. Сомнительно, чтобы некоторые коричневые и лиловые формы отчетливо распознавались собаководами. В любом случае они были бы необычны.
Ген b действует только на черный пигмент, следовательно, все рыжие фенотипы выглядят обычно, но присутствие этого гена выявляется по изменениям окраски радужины глаз, кожи, когтей. Ген d ослабляет рыжий до кремового, при этом изменения пигментации кожи, глаз и когтей незначительны.
Соболиный окрас имеет генотип A
y-B-D-E-. Интенсивность затенения зависит от наличия модифицирующих полигенов. Причем, черное затенение может быть ослаблено до коричневого под воздействием гена b, тогда получится шоколадно-соболиный окрас. Однако, сомнительно, чтобы такой окрас можно было легко распознать. Многие собаководы определяют такой окрас как желтый или рыжий, особенно, если затенение незначительно и тогда оно не выделяется на рыжем фоне. Соответственно, голубо-соболиный или лилово-соболиный сразу и не отличить от простого кремового или палевого.
Аллели белого окраса, альбинизма
Серия аллелей белого окраса или альбинотическая фундаментально контролирует синтез пигмента по всему телу. Можно только удивляться, почему целая серия генов так названа, хотя альбинизм чрезвычайно редок среди собак. Ответ следует искать в общей генетике окрасов млекопитающих. Альбинизм может быть редок у собак, но он обычен у многих млекопитающих. Локус альбинизма более мутабилен, чем другие локусы. Таким образом, появилась серия аллелей с характерными фенотипами. Более того, эти фенотипы сходны у всех видов млекопитающих, поэтому общие знания хорошо применимы и в случаях собак, так как явление альбинизма у них крайне редко.
Общее количество мутантных аллелей локуса альбинизма неизвестно, но, по крайней мере, три из них установлены.
Фенотипическое проявление - Символы гена
Полное проявление окраса — С
Шиншилла — c
ch
Голубоглазый альбинизм — c
b
Полный альбинизм — c
Все нормально окрашенные собаки имеют ген С, который разрешает экспрессию генов синтеза пигментов. Типично, что аллели, следующие ниже в таблице, разрешают экспрессию все меньшего количества пигмента. Последний ген в таблице полностью запрещает синтез любого пигмента.
Аллель шиншилла (c
ch) характерен тем, что осветляет рыжий до бледно-желтого, и почти или совсем не влияет на развитие черного пигмента. Похоже, что этот ген присутствует у пород, имеющих бледно-желтый окрас, таких, как норвежский элькхунд. Ген c
chосветляет как А
у- рыжих; так и ee-рыжих собак. В частности, наиболее ожидаемым генотипом для золотого ретривера следует признать c
chc
chee. Окрас таких шиншилловых животных может варьировать от теплого кремового до почти белого. Большинство этих вариаций образуются под действием модифицирующих руфус-полигенов. Более подробно о них будет сказано ниже. Литтл же предположил, что значительные вариации этого окраса обязаны существованию более чем одного аллеля шиншилла у собак.
Второй аллель шиншиллы, обозначенный Литтлем, как c
e, возможно, отвечает за почти белый фенотип с кремовой тенью вдоль спины, на плечах и голове. Если принять существования этого аллеля, то генотип этих окрасов будет А
у-c
ec
eили c
ec
eee. Можно также предположить, что белые вариации окраса у некоторых пород имеют эти же генотипы, например, вестхайленд-уайт-терьер.
Ни один из полностью белых окрасов у собак не является настоящим альбиносным, так как при этом собаки имеют темные глаза. Следует заметить, что возможность существования аллеля типа шиншилла c
chосновывается более на фенотипических наблюдениях, чем на результатах экспериментальных скрещиваний.
Существование нескольких аллелей шиншиллы можно подвергнуть сомнению, предположив, что не шиншилла, а модифицирующие полигены ослабляют рыжий до палевого во многих случаях. В конечном итоге, пока следует признать достоверным существование только одного аллеля гена шиншилла.
Литтл часто упоминал в своей книге, что ген С не полностью доминантен по отношению к c
ch. Но это противоречит наблюдениям у других видов животных. Однако, если признать, что в случаях, описанных Литтлем, рыжий окрас ослабляется под действием полигенов, а не c
ch, тогда неполное доминирование С не удивительно.
Персон и Ашер (1929) описали два очень светло-окрашенных фенотипа со светло-голубыми глазами и тускло-красными зрачками. Ряд исследователей считают, что, по крайней мере, один из них несет аллель альбинизма, который был назван как «голубоглазый альбинизм» (c
b). О наследуемости c
bочень мало известно. Предложено поместить этот ген между c
chи с в представленной выше таблице.
Полный альбинизм, выраженный в чисто белой шерсти, розовых зрачках и просвечивающей красной радужине, крайне редок у собак. Витни (1947) и Литтл (1957) описали таких альбиносов у пекинесов.
Ослабление окраса, сопряженное с розовоглазием
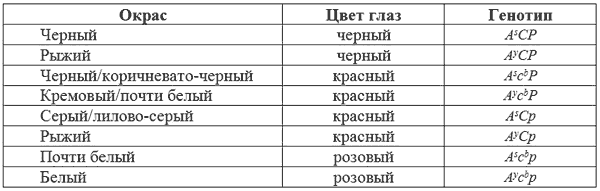
Эта форма ослабления, в целом, обычна для млекопитающих, но редка у собак. Возможно, что одна из форм осветленного фенотипа с голубыми глазами, описанная Персоком и Ашером (см. выше) имела этот ген. Для подобного типа ослабления характерны голубоватый или сероватый окрас и красноватые глаза. Его следует отличать от обычного ослабления до голубого (ген d), когда зрачки остаются темными. Несмотря на уменьшение количества пигмента, «розовоглазое ослабление» не связано с локусом альбинизма, а является мутантной формой (р) гена Р, также отвечающего за развитие интенсивной окраски шерсти и глаз.
При скрещивании между некоторыми из обнаруженных Персоном и Ашером «альбиносов» были получены щенки с темно-окрашенными шерстью и глазами, что может произойти только, если у родителей за окрас отвечали разные независимые гены, продуцирующие розоватые глаза и осветленный окрас шерсти. Например, кроссы между CCpp (розовоглазое ослабление) а c
bc
bPP (голубоглазое ослабление) дают интенсивно окрашенное потомство (Cc
bPp).
Красноватые глаза не всегда полностью лишены пигмента (в противном случае они были бы розовыми) и радужина у них часто голубоватая.
К сожалению, Персон и Ашер описали всех обнаруженных красноглазых собак как альбиносов, хотя не все они таковыми были. Рассмотрим возможные комбинации взаимодействие локусов А, С и Р.
Гены c
bи p взаимодействуют друг с другом в генотипе c
bc
bpp и продуцируют псевдоальбиносов. При этом, каждый действует так, чтобы удалить пигмент, разрешенный другим. Глаза в этом случае должны быть розовыми, а шерсть почти или совершенно белая. У животных, гомозиготных по b (bb) гены c
bи p могут иметь больший эффект на окрас глаз, чем у имеющих В.
Грифельно-серое ослабление
О гене грифельно-серого окраса известно очень мало. Он наследуется как доминантный и фенотипически сходен с геном ослабления до голубого (d). Этот ген обнаружен лишь однажды у колли (символ Sg).
Ослабление по типу «пуховки»
Это необычное ослабление в значительной степени промежуточно. Щенячья шерсть таких мутантных черных особей сначала серая, потом, к 6-8 месяцам, становится нормальной черной с легким, светловатым оттенком. Ген не оказывает воздействия на желтый пигмент и цвет мочки носа. Ген «пухового ослабления» обнаружен пока только у колли (символ pp) (Лунд и др., 1970). Как действует и как наследуется этот ген неизвестно.
Мраморный окрас (мерль)
Мерль — название, данное для обозначения окраса, при котором наблюдается неровное прокрашивание в виде более темных и более светлых участков одного и того же цвета. Такой окрас встречается у шелти, колли, такс, мраморных догов (арлекинов). Окрас мерль получается в случае гетерозиготного состояния гена М, который доминантен по отношению к исходному т. Светлые участки получаются при смешении нормальных и осветленных волос, причем, это более заметно на черном фоне, чем на рыжем. В гомозиготном состоянии ген мерль дает сплошной белый окрас, частично или полностью голубую радужину глаза, часто редуцированное глазное яблоко, частичную или полную стерильность.
Генотипы наиболее распространенных мерлевых окрасов следующие:
Окрас—
Генотип
Голубой мерль — A
s-Mm
Голубой/биколорный мерль — a
ta
tMm
Рыжий/соболиный мерль — A
y-Mm
При «голубом» или пятнистом мерле районы нормальной и голубой окраски резко различаются. В типичном случае — это голубая собака с черными пятнами неправильной формы, разбросанными на голубом фоне. Сходный эффект можно видеть у черно-подпалого мерля, в большей степени на черпаке и в меньшей — на животе. Ген М — больше влияет на черный/коричневый пигмент, нежели на желтый, поэтому, у чисто рыжих собак мерлевая пятнистость не так явно выражена. Замечено, что ген М усиливает белую пятнистость.
Из-за того, что ген мерль в гомозиготном состоянии несет ряд нежелательных признаков, не следует скрещивать гетерозигот, а тем более гомозигот между собой. Логичнее получать мраморных собак, скрещивая мраморных и нормально окрашенных особей. В этом случае получатся как мраморные, так и нормальные щенки.
Возможно, что существует более, чем один аллель гена мерль. Это следует из различий в фенотипическом проявлении и результатов скрещивания (Schaible и Brumuiugh, 1976). Одним из них можно считать ген M
h, обуславливающий мраморный окрас дога (арлекин). Этот ген продуцирует белые пятна в гетерозиготном состоянии (M
hm)- Гетерозиготы M
hM сходны с ММ по фенотипическим проявлениям (белый окрас, частичная глухота, микрофтальмия). Признаки M
hM
h, возможно, также сходны с ММ или даже более выражены.
CN — ослабление
Этот ген ослабляет как черный пигмент, так и желтый: черный до тускло-серого, а желтый до бежевого или почти белого. Соболиный окрас трансформируется в серебристо серый или почти белый в соответствии со степенью затененности. Мочка носа окрашена в светло-рыжевато-коричневый цвет, что заметно отличает действие гена cn от действия других генов ослабления окраса. Волос CN-щенков имеет более тонкое строение и может даже казаться слегка волнистым. Этот эффект исчезает у взрослых. Ген «CN — ослабления» наследуется рецессивно (символ cn) и является полулетальным, вызывая дефицит нейтрофилов крови, что серьезно влияет на способность индивидуума противостоять бактериальной инфекции. Большая часть щенков, несущих cncn погибает в течении нескольких первых месяцев жизни. «CN — ослабление» пока известно только для колли.
Прогрессирующее поседение
Литтл (1957) предположил, что прогрессирующее поседение, или посерение, которое наблюдается у некоторых пород собак, происходит благодаря действию полудоминантного гена G. Автор не приводит экспериментальных данных по скрещиванию для подтверждения своего предположения, однако, Витни (1952, 1958) опубликовал результаты, которые указывают на то, что прогрессирующее поседение детерминировано одним геном. Сначала он обозначил этот ген как si, потом согласился с термином Литтля (G). Характер наследования гена (рецессивный, полудоминантный или доминантный) до сих пор еще не ясен окончательно.
Термин «поседение» употребляется в генетике млекопитающих в случае, когда надо отметить появление значительного количества белых волос на теле. В этом случае количество темных волос превосходит количество белых и животное кажется темным, испещренным белыми волосками. Если количество белых волос больше, чем темных, то такой окрас называют чалым. Непонятно пока, то ли это две отдельные генетические формы, та ли различные степени экспрессии одного гена.
Поседение также классифицируется, как постоянное и прогрессивное. В первом случае оно возникает на определенной стадии развития и остается неизменным с возрастом. Прогрессивное поседение возникает постепенно на протяжении значительного периода или всей жизни. Поседение по второму типу связано с геном G.
Щенки, имеющие ген G рождаются черными (или интенсивно окрашенными), потом сереют (светлеют). Гетерозиготы Gg могут изменяться до темно-серо-голубого. В некоторых случаях это происходит достаточно быстро, но, в основном, животные седеют на протяжении всей жизни. Сходные изменения происходят с гомозиготами, но более резко и глубоко.
Поседение начинается в возрасте нескольких недель и со временем окрас становится серо-голубым. Причем, степень выраженности варьирует как от породы, так и от индивидуума. Поседение может происходить равномерно по всей поверхности туловища, а может и частично. Так, например, у бедлингтона верх головы и часть плеч могут стать почти белыми.
В комбинациях с другими аллелями Gg дает слабое осветление окраса, тогда как GG- резкую редукцию пигмента.
Осветление окраса за счет генотипа GG означает, что ген G если не полностью, то частично эпистатичен по отношению к другим генам окраса. Приведем ниже генотипы, представляющие наибольший интерес:
Окрас—
Генотип
Серебристый — A
s-B-D-E-GG
Шоколадно-серебристый — A
s-bbD-E-GG
Голубо-серебристый — A
s-B-ddE-GG
Кремово-серебристый — A
s-B-D-eeGG
Светло кремовый, серебристый — A
s-B-ddeeGG
Животные имеющие вышеперечисленные окрасы являются серебристыми или чалыми, у которых белые или слабопрокрашенные волосы перемежаются с окрашенными. При этом, если на тела много окрашенных волос, то цвет их легко фенотипически идентифицировать, но, если их мало, определить окрас можно лишь по общему впечатлению. Так, все bb можно идентифицировать по наличию коричневой мочки носа и осветленных глаз. Шерсть может быть с легким коричневатым оттенком. Таким же образом ee-серебристые животные могут иметь бледно-желтый или кремовый оттенок. Все фенотипы можно легко распознать до начала поседения. Следует отметить, что A
s-B-ddE-GG должны иметь голубой окрас даже до начала поседения благодаря наличию гена d, а не G. Экспрессия гена G может быть не так заметна благодаря аффекту ослабления за счет гена d. Любая порода, где какие-то особи рождаются интенсивно окрашенными, а затем постепенно бледнеют до отчетливо более светлого окраса, имеет в своем генофонде ген G. Очевидно, что он имеется у таких пород, как бедлингтон, керриблютерьер, бобтейл, пудель. Литтл (1957) предложил, что ген G может присутствовать у кернтерьера, денди-динмонт-терьера, скайтерьера и йоркширского терьера. Можно также предположить его наличие у австралийского терьера и австралийского шелкового терьера.
Белая пятнистость
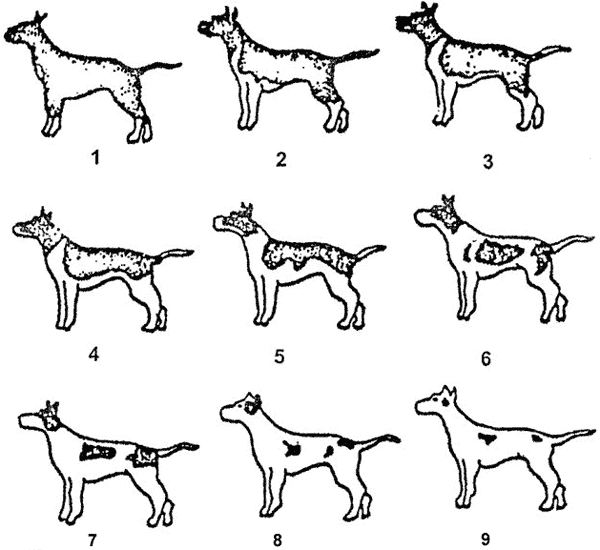
Наличие белого узора на туловище характерно для многих пород собак. Более того, у многих из них характер расположения белых отметин является чуть ли не основным признаком. Теоретически, белые участки рассматриваются как белые пятна, не взирая на количество белого. Раньше этот термин употреблялся только, когда количество белого лимитировано в виде пятен на темном фоне. Теперь термин «белая пятнистость» употребляется более широко и включает также случаи, когда белый преобладает или же собака почти вся белая. При этом белое поле можно рассматривать как одно большое белое пятно.
Распределение и распространение белых отметин следует относительно правильной прогрессии. Так, если расположить пятнистых собак по мере убывания прокрашенных участков, то выявляется следующая закономерность (см. рис. на стр. 87). Сначала белые отметины появляются на груди, лапах, морде и кончике хвоста. По мере увеличения площади белый распространяется на всю грудь, живот, конечности. Долее — на плечи и холку, затем бока и верхнюю часть спины. Постепенно, на теле остаются немногочисленные пигментированные пятна, уменьшающиеся в размерах. На этой стадии также наблюдается закономерность в расположении оставшихся окрашенных пятен. Чаще всего они остаются на боках, голове, особенно вокруг ушей и в основании хвоста. На рисунке схематично отображены описанные процессы.
Причиной такой общей закономерности развития белой пятнистости состоит в отсутствии пигмент-образующих клеток в коже раннего эмбриона при корнях волос. Эти клетки образуются в определенных местах, что соответствует приблизительно участкам на голове и вдоль спины у развивающихся эмбрионов. Если что-то препятствует образованию достаточного количества первичных точек пигментации или скорость распространения пигмент-образующих клеток по туловищу тормозится в ходе развития эмбриона, то щенки рождаются с белыми отметинами. Хотя структура кожи остается нормальной и на ней вырастают нормальные волосы, они будут бесцветными (т.е. белыми для человеческого глаза), так как пигмент-продуцирующие клетки не успели распространиться в эти участки. Так как эмбриональное развитие — это строго детерминированный процесс, то для этих клеток возможность своевременно занять свою позицию так же строго ограничена во времени.
Следует, однако, отметить, что, несмотря на общие тенденции в увеличении количества белого, существуют значительные вариации. Особенно это касается собак с преобладанием белого. У них пятна могут возникать в любом месте головы, плеч, боков и, как правило, не обязательно симметрично. Напротив, положение белой проточины на морде и отметин, образующих воротник (так называемая воротниковая степень пятнистости) относительно постоянно и служит отличительной чертой таких пород, как бостон-терьер, колли и сенбернар.
Животные, у которых на белом фоне разбросаны пигментированные пятна, называются пегими. Некоторые из этих вариаций обусловлены действием полигенов, модифицирующих экспрессию (т.е. количество белого) основных аллелей пятнистости, а некоторые — нарушением в ходе эмбрионального развития.
Показано, что, если в породе предпочтителен какой-то определенный характер пятнистости, то количество других вариаций значительно уменьшается, но не исчезает совсем.
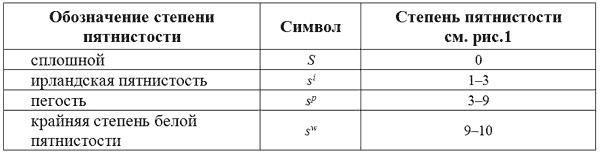
Литтл (1957) выдвинул гипотезу, что разнообразие распределения пятен на туловище можно объяснить существованием трех мутантных аллелей гена S, определяющего сплошной (небелый) окрас.
Аллель ирландской пятнистости продуцирует наименьшую степень пятнистости. Белые отметины варьируют от пятен начиная с морды, груди, плеч, головы, ног и кончая узким воротником на шее. Типичным примером экспрессии вариабельности ирландской пятнистости s
is
iпо Литтлю является бассенджи.
Животные, гомозиготные по гену пегости s
ps
p, имеют большее количество белого, чем животные предыдущего типа. Однако, для s
pхарактерно значительно более широкая вариабельность экспрессии. Например, по Литтлю, бигль имеет генотип s
ps
p, но значительная часть белых отметин по характеру распределения лишь ненамного отличается от бассенджи. С «другой стороны фокстерьеры имеют такой же генотип s
ps
p, но в общей своей массе имеют столько белого, что порой их можно рассматривать как белых собак с небольшими пигментированными пятнами.
Аллель крайней степени пятнистости определяет наибольшее количество белого и, в крайнем случае, такие животные полностью белые за исключением небольшого пятна или пятен на крупе или голове. В эту группу также входят темноглазые собаки совсем без пятен. У целого ряда пород такой фенотип является или стандартным окрасом или принятой вариацией.
Доказательством того, что чисто белые темноглазые собаки имеют генотип s
ws
wявляется тот факт, что у таких собак могут иногда появляться щенки с небольшими пятнами в стандартных местах. Поэтому, Литтл предположил, что белые бультерьеры, большие пиренейские собаки, самоеды и салихемтерьеры имеют генотип s
ws
w. Возможно, что аллели пятнистости не полностью доминантны друг к другу, но эти взаимоотношения неустойчивы.
Исходя из многих наблюдений, можно предположить, что ген S доминантен к s
i, но не к s
pили s
w. Гетерозиготы Ss
pи Ss
wдают окрасы по типу ирландской пятнистости. Аллель s
iдоминантен к s
p, но не к s
wГетерозиготы s
is
wдают пегость или окрас, близкий к этому. Взаимоотношения между s
pи s
wпока не ясны из-за недостатка данных. Возможно, что s
ps
wдают белых собак с незначительным количеством окрашенных пятен. Следует помнить, что экспрессия s
pпоказывает огромную вариабельность и серьезно усложняет окончательное выявление взаимоотношений между аллелями этого локуса.
Кроме этого, экспрессия всех мутантных аллелей пятнистости частично контролируется модифицирующими полигенами. В большей мере они определяют независимо от породы степень пятнистости, нежели порядок, местоположение пятен и их конфигурацию.
До сих пор не выяснены механизмы действия модификаторов: то ли они действуют сами по себе, то ли влияют на экспрессию S-аллелей. Почти очевидно, что некоторые из них способны сами по себе продуцировать начальную степень белой пятнистости у гомозиготных SS (генотипически — это генотип сплошного окраса). В некоторых случаях небольшие белые пятна могут возникать как следствие сбоев эмбрионального развития. Однако, сомнительно, чтобы все подобные случаи можно было бы объяснить таким образом. Ситуация усложнена еще и тем, что многие полигены, вовлеченные в этот процесс, экспрессируются также вариабельно. Таким образом, многие SS-животные, имеют незначительные белые отметины за счет действия модификаторов. В любом случае, при постоянном отборе только полностью пигментированных собак, всегда будут случаи появления небольшой части животных с белыми отметинами.
И Витни (1947) и Литтл (1957) отмечали, что черные и красные спаниели отличаются по количеству белого. Так, у большей части черных собак наблюдается небольшое или среднее количество белого, тогда как у рыжих спаниелей белого значительно больше. Можно предположить, что один из наиболее влиятельных модификаторов гена s сцеплен с геном е, или, что пигмент-продуцирующие клетки, содержащие в геноме ее менее способны распространяться по коже сравнительно с E-содержащими. Такие различия не проявляются у полностью окрашенных животных, но становятся очевидными у пятнистых. Берне и Фразер (1966) отметили, что коричневые собаки занимают в этом отношении промежуточное положение. Это подразумевает, что ген b является фактором, также влияющим на развитие пятнистости.
Может показаться странным, что различные гены белой пятнистости являются аллелями одного локуса, так как у многих видов млекопитающих белая пятнистость возникает при мутациях различных локусов. Сходную ситуацию можно было бы предположить и для собак, как отмечает Хатт (1979). Однако, обширные данные, опубликованные Литтлем (1957) полностью согласуются с предположением о существовании только одного локуса с несколькими аллелями. Существование независимого от s-аллелей гена белой пятнистости сомнительно. Вопрос лишь в том, сколько на самом деле аллелей: две или три, так как очень трудно порой различить экспрессию s
iи s
p.
Тиковость (тиковая пятнистость, крапчатость)
У многих пород, таких, как пойнтеры, сеттеры и спаниели, белые участки покрыты многочисленными пигментированными мелкими пятнами. Такое явление называется тиковой пятнистостью и определяется геном Т. Тиковая пятнистость отсутствует при рождении и проявляется в возрасте от нескольких недель жизни щенка до первой линьки. Экспрессия гена Т очень вариабельна: от нескольких небольших пятнышек до такого их изобилия, что белые участки тела кажутся чалыми. У длинношерстных пород тиковость выглядит как чалость благодаря тому, что длинные волосы разных цветов легко перемешиваются. У некоторых пород наблюдается тенденция к усилию тиковости на ногах и животе.
Тиковость является результатом взаимодействия между любой из аллелей белой пятнистости и гена Т. Чаще всего это аллель s
pкак наиболее распространенный и продуцирующий белые участки достаточной протяженности для четкого проявления тиковости. Тиковость не влияет на гены окраса, т.е. цвет крапа соответствует основному окрасу. Генотип особей с тиковой пятнистостью должен быть s
ps
pT-, s
ps
wT- или s
ws
wT-. При этом следует помнить, что количество белого очень вариабельно, и, что тиковость проявляется только на белом фоне. Остальная часть генотипа, определяющего окрас, будет соответствовать цветовой вариации особи. Например, тиковый черный имеет генотип A
s-B-D-E-s
p-T-, а рыжий тиковый A
y-B-D-E-s
p-T-. Черно-подпалые тиковые (a
la
lB-D-E-s
p-T-) имеют черный крап на спине и рыжий на животе.
Раньше считали, что пятнистость по типу далматинов определяется отдельным геном, но Литтл (1957) убедительно показал, что это модификация обычной тиковости. Например, черные далматины имеют генотип A
s-B-D-E-s
ws
wT-. Разведенцы далматинов вывели собак с более крупными, яркими, прокрашенными пятнами, случайно разбросанными по всему телу.
Экспрессия гена тиковой пятнистости Т варьирует от слегка крапчатые собак с несколькими крапивами только на конечностях и животе до сильно крапчатых животных, часто встречающихся у гончих пород (например, гасконская гончая и гриффон). В последнем случае собаку с таким окрасом можно назвать чалой. С другой стороны, варьирует не только количество крапа, но и качество его. Так что тиковость является весьма вариабельным признаком. Является ли это результатом отбора и закрепления в генотипе определенного количества полигенов, управляющих экспрессией гена Т, еще предстоит выяснить. Литтл заметил, что при скрещивании далматинов и собак с окрасом, похожим на далматинский, в первом поколении происходит возврат к обычной крапчатости. Возможно, что существует не один ген тиковости, каждый со своим характерным типом крапчатости. Однако, тот факт, что у собак наблюдается экстраординарная вариабельность экспрессии тиковости сам по себе не может служить этому доказательством.
Маски
Маска — термин, употребляющийся для обозначения черного поля на морде, часто покрывающего переднюю часть «лица» и ушей. Наиболее четко маска видна у А
у-рыжих собак, например, у таких разных пород, как желто-коричневая (оленья) вариация гигантского мастифа и у миниатюрного мопса.
У чепрачных и черно-подпалых собак маска, если она есть, затемняет подпалы, ассоциированные с генами a
saи a
t. В этом случае получаются черномордые чепрачные и черно-подпалые собаки. У первых, ген маски трудно выявить, т.к.
наличие или отсутствие темной морды можно также рассматривать и как вариации чепрачности. Черно-подпалый окрас более стабилен и однообразен, поэтому маска у черноподпалых собак сразу же обнаруживается, благодаря отсутствию подпалов на морде. Естественно, что маску невозможно обнаружить у черных A
s-собак, хотя в геноме соответствующий ген может присутствовать.
Маска может варьировать по степени развития. Так, она может покрывать всю «лицевую» часть головы и уши. Но это не обязательно означает влияние полигенов. О характере наследуемости маски известно очень мало. Литтл (1957) ссылается на случай рождения от рыжей таксы и мопса оленьего окраса с маской всех восьми щенков с черными масками. Это указывает на моногенность и доминантность маски как признака. Условно, ген маски можно обозначить как Ма.
Литтл считает, что существует несколько аллельных генов маски, но не дает четких доказательств. Тигровые животные могут быть как о маской так и без нее, что само по себе достаточно, чтобы отбросить всякие сомнения о моногенности признака. Геи маски, возможно, не экспрессируется у ee-животных, но и это не свидетельствует о наличии серии аллелей. Возможно, что ee-рыжие животные не могут вообще синтезировать черный пигмент в волосе. Отсутствие маски у ee-рыжих собак можно рассматривать как признак, отличающий их от А
у-рыжих животных. Все рыжие собаки с маской имеют ген А
у, тогда как рыжие без маски могут иметь как Ay так и ee, но, скорее всего последний.
Руфизм (рыжина)
Интенсивность желтого пигмента шерсти может варьировать от бледно-кремового до насыщенного рыжего. Некоторые из вариаций можно рассматривать как результат действия основных генов окраса, таких как d или, возможно, c
ch(шиншилла), но не все. Остается еще значительное количество вариаций, которые обязаны действию группы полигенов, известных как руфус-полигены. Причем, эти полигены воздействуют как на A
yтак и на ее. Интенсивность пигментации варьирует в зависимости от особи, линии или породы. Эти же модификаторы также определяют интенсивность пигментации подпалов у чепрачных и черно-подпалых собак, у которых они могут варьировать от бледно-желтых до насыщенно красных. Это поднимает статус данной группы генов о простых модификаторов генов А
уи е до фундаментальной группы генов, определяющих основной составной элемент окраса. Только сплошной черный или сплошной белый окрасы не подчинены влиянию руфус-полигенов.
Действие этих генов в основном направлено на усиление пигментации. Волк имеет тусклый, однообразный окрас и собаководы прошлого, очевидно, отбирали более яркоокрашенных особей, определяя таким образом внешний вид большинства пород. Ильин (1932, 1941) обнаружил, что более бледный окрас доминантен по отношению к более яркому и объяснил это существованием серии аллелей «интенсивности» (символы Int, int
mи int). Аллель Int продуцирует наиболее бледные тона, a int — наиболее яркие, int
mзанимает промежуточное положение. Вполне возможно, что интенсивность желтой пигментации действительно регулируется мажорными генами (а не полигенами), но, чтобы доказать это, у Ильина было слишком мало наблюдений. Скорее всего, в этот процесс вовлечены многочисленные руфус-полигены. Наблюдения Ильина интересны и, возможно, полезны, т.к. потерю яркого рыжего окраса у гибридов между яркими и осветленными собаками наблюдали Njebbes и Wriedt (1927). Бернс и Фразер (1966), а также Виллис (1976) заметили, что насыщенность рыжего окраса может быть легко потеряна и трудно затем восстанавливается. Это означает, что, если есть усиливающие и ослабляющие полигены, то последние должны быть доминантны по отношению к первым. Однако, для этой группы полигенов характерно, что те, которые доминантны в одних скрещиваниях, становятся рецессивны в других, в зависимости от родителей. Т.о. очень трудно выявить какие-либо закономерности, лежащие в основе взаимодействия этих генов.
Вариабельность чепрачности
Экспрессия гена чепрачности настолько вариабельна, что можно предположить наличие более, чем одного аллеля, определяющего этот признак. Например, один аллель дает темно-чепрачный окрас, другой — светло-чепрачный. Вариации чепрачности составляют непрерывный ряд, что можно объяснить скорее влиянием модифицирующих полигенов. Чепрачный окрас часто можно спутать с соболиным, т.к. он имеет тенденцию с возрастом осветляться. Это говорит о генетической близости аллелей a
saи A
y. С другой стороны, когда чепрак распространяется на большую часть тела, окрас становится похожим на черно-подпалый.
В идеале, чепрачными принято называть собак с V-образным темным пятном на спине и боках туловища, простирающимся от хребта до живота. Однако, как распространение, так и интенсивность пигментации этой области может варьировать. При яркой, интенсивной пигментации чепрачный окрас легко идентифицировать, но при более светлой пигментации границы чепрака размыты и наличие его трудно распознать. Фактически, этот окрас стремится иметь сходство с диким волчеобразным. И это является основой для предположения, что ген чепрачности является модификацией исходного гена А. Сходство усиливается, если рыжий подпал осветлен. Строго говоря, вариабельность чепрачности определяется двумя независимыми системами модификаторов. С одной стороны — это модификаторы, управляющие вариабельностью чепрака, его интенсивностью, формой и протяженностью. С другой стороны, окрас подпалов определяется руфус-полигенами, как в случае с чисто рыжими собаками. Причем, они не влияют на экспрессию гена чепрачности.
Интересно было бы предположить, что некоторые фенотипические вариации чепрачного окраса обязаны полигенам, влияющим на экспрессию гена А, которые затеняют рыжий окрас превращая его в соболиный. Такие затеняющие полигены могли бы продуцировать некоторые вариации чепрачного окраса, модифицируя экспрессию гена А. К сожалению, никакими экспериментальными данными по этому вопросу мы не располагаем. Интенсивность пигментации подпалов может также варьировать, причем независимо от интенсивности окраса чепрака. Почти наверняка это происходит благодаря действию независимых руфус-полигенов.
Полигены затенения или соболиного окраса
Эти полигены, судя по названию, вызывает потемнение шерсти. Их наиболее очевидной экспрессией является измелен не обычного А
у-рыжего окраса до соболиного. Большинство, если не все А
у-рыжие особи имеют некоторое количество черно-типированных волосков, как правило, на голове и вдоль спины. Туловище щенка обычно покрыто темными волосами, придавая ему более темный окрас. При взрослении щенка окрас становится все более светлым, пока не будет достигнут окрас взрослого животного.
Когда количество темных волос многочисленно у взрослого животного рыжего окраса, получается соболиный окрас. Степень затененности значительно варьирует, особенно на голове, плечах и спине. Наиболее темные особи могут напоминать осветленных чепрачных. Особенно, если затенение распространяется на бока, напоминая по форме сглаженный чепрак.
Типичный соболиный окрас можно встретить у колли или шотландской овчарки. Показано, что только А
у-рыжий окрас конвертируется в соболиный с помощью полигенов затенения. Предполагается, но еще не доказано, что эти полигены не оказывают сходного воздействия на ген е, который, очевидно, эпистатичен по отношению к данным модификаторам.
Белые собаки
Фенотипически, полностью белые собаки могут быть основой для ряда пород или породных вариаций. В принципе, полностью белые собаки имеют различную генотипическую основу. У многих млекопитающих встречается альбинизм. Этот фенотип полностью лишен Пигмента не только в волосах, но и в радужине глаза, кожи, когтях и др. Такие белые особи красноглазы. Однако, когда собаководы говорят о белых собаках, они имеют в виду темноглазых животных. Таких собак нельзя назвать альбиносами, даже если они имеют голубые глаза.
Одно из наиболее ранних предположений о происхождении чисто белых собак состоит в том, что окрас таких особей есть следствие действия гена крайней степени пятнистости. Литтл постулировал существование аллеля s
w, который и обуславливает чисто белый окрас. Животные с генотипом s
ws
wили полностью белые, или имеют незначительные отметины в области ушей и основании хвоста. Постоянная селекция и удаление особей с отметинами из разведения в прошлом привели к тому, что они составляют небольшую часть популяции. Однако, факт, что такие особи все-таки появляются, является хорошим подтверждением существования генотипа s
ws
w. В результате, Литтл смог предположить, что у бультерьера, большой пиренейской собаки, самоеда и силихэмтерьера имеет место быть генотип s
ws
w.
Можно также предположить, что чисто белый окрас возникает в результате действия гена шиншилла (c
ch), который удаляет почти весь желтый пигмент волос, но не глаз. В результате могут получиться темноглазые белые собаки. Tjebbes и Wriedt (1927) предположили, что такими собаками могут быть самоеды. Возможно, что они ошибались, исходя из более поздней концепции Литтля. Однако, совсем не обязательно, что все самоеды несут s
ws
w, т.к. данные Литтля получены при исследовании американской популяции самоедов, а Tjebbes и Wriedt наблюдали скандинавскую группу.
Как известно, пигменты в пигментных гранулах могут быть только двух видов: желтый и черный. Синтез черного пигмента запрещают два гена — гены А
уи е. Это первый шаг к возникновению белого окраса. Для получения белой собаки нужен еще один ген, который удалял бы уже желтый пигмент. Этим геном является c
ch, т.к. именно он, главным образам, запрещает синтез желтого пигмента.
Хотя ген c
chописан Литтлем (1957), существование его не бесспорно. На данный момент мы допускаем, что он есть и действует только в отношении желтого пигмента, оставляя черный. Тогда, комбинация A
yс c
chпродуцирует белый окрас, т.к. A
yудаляет черный, a c
chудаляет оставшийся желтый. Ген А
уиногда не полностью удаляет черный пигмент. Поэтому, возможно получить затененный белый или белый с маской. Ген е более эффективно удаляет черный пигмент. Следовательно, более чистобелые собаки будут скорее c
chc
chee, чем A
y-c
chc
ch.
Однако, c
chможет недостаточно эффективно удалять желтый пигмент. Такие животные могут быть сходны с золотым ретривером бледно-кремового окраса. Поэтому, требуется дополнительная селекция для удаления остатков желтого пигмента и получения чисто-белых собак.
Осветление и фактическое исчезновение желтого пигмента может быть зависимо от осветляющих руфус-полигенов. Белые животные такой категории могут быть отличимы, если они дают собак с кремовым оттенком по всему корпусу, особенно вдоль позвоночника, или получаются от бледно-кремовых родителей. Литтл (1957) считает, что вестхайлендвайт-терьер может служить примером белого окраса, обусловленного генотипом A
y-c
chc
c
h.
Ген мраморного окраса М в гомозиготном состоянии дает белый или почти белый окрас с небольшими участками окрашенных волос на голове. Некоторые индивидуумы развиваются в прекрасных собак, но большая часть имеет один или несколько дефектов. Сюда можно отнести глухоту, редукцию глазного яблока, вплоть до полного его отсутствия. В целом, очевидно, что белые животные, имеющие ген мерль (М) не должны скрещиваться.
Ильин (1932) сообщает, что некоторые белые вариации собак обязаны доминантному гену W. Однако, Витни (1947) утверждает, что в его экспериментах на белых собаках ген W ведет себя как рецессивный. Вполне возможно, что они несли ген s
w.
Окрас радужины глаза
Под цветом глаз обычно подразумевается цвет радужины, т.к. зрачок у большинства собак черный. Как правило, радужина у собак коричневая, интенсивность пигментации которой, варьирует. Глаза могут быть также ореховые или янтарные. Цвет радужины определяется двумя факторами: действием основного гена, который определяет основной цвет и действием модификаторов, которые создают дополнительное разнообразие цветовых вариаций радужины.
Кроме этого, ген коричневого окраса b и ослабитель d также влияют на цвет радужины. У bb-животных она светлее, чем у В-. Такой цвет называют ореховым. В свою очередь, интенсивность его варьирует под действием полигенов. Ослабитель d осветляет глаза до янтарного или даже желтого.
Бернс (1943) описывает радужину голубых собак как дымчатую или дымчато-желтую.
Он также отметил, что цвет глаз обычно окрашенных собак варьирует от темно- до светло-коричневых, и заметил, что собаки со светло-коричневыми глазами при скрещивании не дают щенков с темно-коричневыми глазами. Напротив, от собак с темными глазами можно получить светлоглазых щенков. Однако, утверждать о моногенности этого признака преждевременно. Более очевидно, что интенсивность пигментации детерминирована полигонами, увеличивающими или уменьшающими количество пигмента в радужине. Те же полигены, вероятно, модифицируют пигментацию глаз и у шоколадных и у голубых собак независимо от действия основных генов d и b, вызывая то разнообразие оттенков, которое мы наблюдаем у собак этих окрасов.
Вариации цвета радужины, описанные выше, присущи целому ряду обычно окрашенных собак. Однако, существуют случаи, о которых следует сказать особо, Некоторые, наиболее крайне выраженные альбинотические аллели, могут давать светло-голубые и даже белесые глаза благодаря резкому уменьшению пигмента в радужине. Происходит это в результате смешения цветов крови и остатка пигмента. В случае полного альбинизма радужина становится бесцветной и сквозь нее просвечивает красная кровь сосудов глаза.
Ген мерль (М) может давать голубую радужину, как одно из своих проявлений. Радужина может быть частично или полностью голубой. Редукция пигмента может также касаться как одного, так и обоих глаз. В таком случае собака вырастает разноглазой. Эффект редукции пигмента глаза находится в прямой связи с ослабляющим действием гена М на окрас шерсти, т.к. экспрессивность его варьирует.
Ильин (1932) приводит примеры голубоглазости, которые не зависят от основного окраса. Описанный им признак ведет себя рецессивно по отношению к коричневоглазости. Витни (1947) отмечает сходные случаи у пуделей. Он сообщает, что голубоглазые пуделя могут появляться и у темно- и у светло-коричневоглазых индивидуумов и подтверждает рецессивность этого признака.
Ильиным были описаны случаи, когда нормальное желто-зеленое свечение в темноте помещалось на красное, что являлось рецессивным по отношению к желто-зеленому.
Цвет мочки носа
В противоположность цвету глаз, окрас мочки носа почти полностью предопределен основными мажорными генами окраса. Интенсивно прокрашенная собака с хорошим здоровьем имеет черную мочку носа. С другой стороны, коричневые животные (bb) имеют коричневую (печеночную) мочку носа, а ослабление по гену d дает асфальтово-серую. Можно наблюдать различные вариации в интенсивности прокрашивания, но, в целом, цвет мочки носа соответствует окрасу животного. Даже в случае красного или желтого окрасов В- или bb- животные могут быть легко различимы по цвету мочки носа.
КАЧЕСТВО ШЕРСТИ
У собак существует значительное количество различных типов шерсти. Однако, различия эти в некоторых случаях скорее видимы, чем реальны. Генов, определяющих качество шерсти может быть всего несколько, а вариации в их экспрессии определяются соответствующими модифицирующими генами. Проиллюстрируем это на примере такого признака, как длинная шерсть. Нет причин считать, что в проявлении этого признака участвует более, чем один ген. Несмотря на это, очевидно, что длинношерстные животные различаются от породы к породе. Эти вариации происходят благодаря полигенам модификации экспрессии гена длинношерстности. Различия в степени длинношерстности закреплены в породах в результате длительного периода их становления.
Для проведения корректного анализа наследуемости длины шерсти следовало бы изолировать различные компоненты качества шерсти и изучать наследственность каждого из них отдельно. К сожалению, в этом направлении сделано крайне мало. Даже у собак с обычным шерстным покровом существует удивительное разнообразие длины волоса, плотности шерсти (количество волос на единицу площади кожи), соотношения числа остевых волос и подшерстка, прямых и имеющих тенденцию к волнистости. Возможно, что перечислены еще не все слагаемые качества шерсти. Многие из них варьируют независимо друг от друга и комбинации их дают большинство типов шерсти.
Т.о., пока не доказано обратное, наследуемость качества шерсти следует считать полигенной.
Длинная шерсть
Длинная шерсть определяется мутантным геном l, возможно, что этот ген продлевает рост волоса, и он становится в несколько раз длиннее, чем обычный. Экспрессия l модифицирована по ряду показателей. У некоторых шерсть не кажется очень длинной и плотно прилегает к телу. На экспрессию гена l указывает в этом случае длинный украшающий рост волос на конечностях и хвосте, как у английского сеттера, у других шерсть особенно длинная, прямая и шелковистая как у йоркширского терьера. У третьих, она обильная и очень плотная, как у чау-чау. Наконец, у последних она длинная, свалявшаяся и косматая, как у бобтейла. У спаниелей длинная шерсть часто имеет волнистую структуру и сильно развита на конечностях. Особенности наследуемости украшающего волоса будут обсуждаться ниже.
Жесткая шерсть
Жесткая шерсть, характерна для эрдельтерьера, таксы, фокстерьера, брюссельского гриффона и дратхаара. Она образуется в результате действия доминантного гена Wh (Whitney, 1947; Winge, 1950). Считается, что у всех жесткошерстных пород работает один и тот же ген, хотя предположение о том, что их больше, можно принять как рабочую гипотезу. Есть наблюдения, что ген Wh задерживает рост шерсти, тогда генотип llWh- дает волосы длиннее, чем LLWh-, но короче, чем llwhwh.
Скрученная шерсть
Витни предположил, что курчавая или скрученная шерсть ирландского водяного спаниеля наследуется рецессивно по отношению к прямой. Условно, ген обозначен k.
Курчавая шерсть
Считалось, что курчавая шерсть ретривера является короткошерстной вариацией скрученной шерсти ирландского водяного спаниеля. Бернс и Фразер (1966) сообщают, что при вязке курчавого ретривера с дратхааром щенки имели такую же курчавую шерсть, как у ретривера. Следовательно, курчавая шерсть, возможно, определяется разными генами у водяного спаниеля и ретривера.
Волнистая шерсть
Витни описал особенности некоторых скрещиваний, которые указывают на то, что волнистая шерсть кокер-спаниелей и ряда других пород, возможно, наследуется рецессивно по отношению к прямой. Однако, это не более, чем предположение. Сведений для прямых доказательств недостаточно, поэтому, предполагаемый ген может быть предварительно обозначен, как wa.
Шерсть с завитком
Пиллингом была описана линия кокер-спаниеля с необычными кольцами волос на плечах и на боках. Некоторые собаки имели, лишь одно кольцо, которое могло располагаться на голове или шее. Этот признак наследовался рецессивно и ген, его определяющий, обозначен как wo.
«Складчатая» шерсть
Витни описал необычную аномалию кожи (в виде складок) или направления роста волос у щенков бладхаунда. В возрасте 1-7 дней у них появлялись длинные, регулярно расположенные волосы, растущие от шеи до задних конечностей. В семь дней они исчезают, и шерсть по всему телу становится нормальной. Результаты скрещиваний позволяют предположить, что этот признак наследуется рецессивно. Ген, кодирующий его обозначен как rp.
Украшающая шерсть
Украшающая шерсть или кайма из длинных волос на конечностях и нижней поверхности хвоста является характерной чертой длинношерстных собак. Степень развития этого признака сильно варьирует. Битнер (1947) сообщает, что кроссы между ирландским водяным спаниелем (с незначительным развитием украшающего волоса на хвосте) и кокером или ирландским сеттером (с обильной оброслостью) давали потомство с обильной оброслостью. Это указывает на доминантность признака, но недостаточно для того, чтобы ответить на вопрос, кодируется ли этот признак одним независимым геном или развивается в результате воздействия модифицирующих полигенов на ген длинношерстности. Последнее наиболее вероятно и может служить примером того, как экспрессия гена длинношерстности может быть модифицирована другими генами.
Особенность роста волос разного цвета
Витни (1947) отмечает, что у длинношерстных, пятнистых собак черные и белые волосы разной длины. У кокера и английского сеттера черные волосы длиннее, чем белые. Черные волосы также длиннее, чем рыжие. Бернс и Фразер (1966) наблюдали подобное и добавляют, что белые волосы в отдельных случаях могут быть той же длины, что и черные, но мягче по структуре. Рыжие волосы также мягче и шелковистее, чем черные.
Бесшерстность
Бесшерстность — это явная аномалия. Однако этот признак следует рассмотреть в настоящей работе, т.к. он является определяющим для таких пород, как китайская или мексиканская голые собаки. Эти животные не всегда полностью бесшерстны, а имеют варьирующее количество волос на голове и конечностях. Летард (1930) показал, что эти породы образовались в результате действия доминантного гена Hr, который в гомозиготном состоянии летален. Следовательно, все бесшерстные особи этих пород гетерозиготны по Hr (Hr hr).
Описаны случаи, когда бесшерстность наследовалась рецессивно. Поэтому, не стоит считать, что этот признак всегда кодируется доминантным геном Hr.
Бесшерстные мутанты время от времени встречаются среди нормальных собак и некоторые из них могут выживать в более теплом климате или специально отбираться. Если такие животные имеют древнее происхождение, то могут приобрести ауру респектабельности и быть признанными как порода. Насколько это оправдано является спорным вопросом.
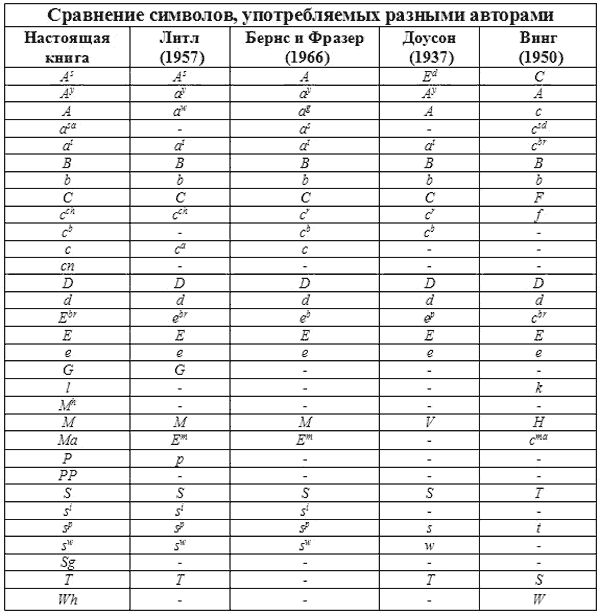
СРАВНИТЕЛЬНАЯ СИМВОЛИКА ГЕНОВ
Читатели, которые интересуются литературой по генетике, рано или поздно сталкиваются с проблемой путаницы в обозначениях генов. Дело в том, что различные авторы пользуются различными символами для обозначения одного и того же гена. Это прискорбно, но неизбежно. Исследователи сразу же не согласовали стандартную номенклатуру генов и называли новые описанные гены как им заблагорассудится. Настоящий раздел книги является попыткой свести накопленное в единую систему.
Впервые на это обратил внимание Доусон (1937), который попытался привести в соответствие предыдущие достижения. Собранный им материал выдержал проверку временем. Последующий вклад Литтля (1957), а также Бернса и Фразера (1966) укладывался в основное русло генетики млекопитающих. Увы, это не относится к работам Винга (1951). Его система полностью противоречит предшествующим и достаточно запутана. Он предлагает ряд генов, в которых нет необходимости. Некоторые гены Винг разместил не в тех аллельных сериях, где следовало бы.
Таблица 11
Таблица 12
Заключение
Если сравнить окрасы собак, обусловленные генами Ay и е, можно заметить, что в первом случае собаки более интенсивно пигментированы. Однако, не стоит считать, что все красные собаки несут Ay, а все более бледные — ее. Там где возникают сомнения по поводу происхождения окраса можно допустить, что он скорее обусловлен А
у, чем ее. Т.е., прежде, чем допустить, что окрас обусловлен генотипом ее, надо доказать, что он не обусловлен геном А
у.
Там, где белая пятнистость является одним из породных признаков, следует учитывать три аллеля, установленных Литтлем (1957). Однако, у многих пород минимальная белая пятнистость является нежелательной или даже запрещенной. Такие отметины возникают чаще всего на морде, копчике хвоста, груди, животе, реже на лапах. Поседение подбородка у многих пород также относится к минимальной белой пятнистости. Сомнительно, чтобы такое многообразие признака кодировались одним геном. Если это так, то этот ген часто изменяет экспрессию самого себя. Это маловероятно, и скорее всего минимальная пятнистость является полигенным признаком, с вариабельной экспрессией. Постоянная выбраковка пятнистых животных означает лишь усиление контроля, но не полная элиминация этого признака. У некоторых пород белые отметины не являются дисквалифицирующим признаком, но нежелательны. Такое послабление понятно, но оно накладывает бремя ответственности, до какой степени допускать проявление этого признака в породе.
Генетическое обоснование качества шерсти является целой проблемой. К настоящему времени четко установлены лишь несколько генов, определяющих структуру шерсти. Наиболее четко можно идентифицировать гены длинной и короткой шерсти. Несмотря на то, что длинношерстность определяется одним геном l, его экспрессия отличается у многих пород (Витни, 1974).
То же самое нельзя сказать о жесткой, курчавой, волнистой шерсти, шерсти с изломом, с завитком и т.д. Возможно, что у разных пород собак каждый из этих признаков контролируется разными генами. Например, у домашней кошки для шерсти с изломом (порода рекс) идентифицировано несколько генов, хотя фенотипическое проявление их очень сходно. С другой стороны, один и тот же ген в разных породах экспрессируется в разной степени, что может привести к ошибочному мнению о существовании разных аллельных вариантов.
В любом случае, при принятии окончательного решения очень легко ошибиться. Поэтому, любая информация, которая бы пролила дополнительный свет на наследуемость многих признаков была бы очень полезна.
Аффенпинчер
В основном, это черная собака с длинной жесткой шерстью. Встречаются также черно-подпалые, соболиные различных оттенков. В соответствии с Литтлем, у некоторых особей имеется ген маски.
Окрас—
Генотип Черный — A
s-llWh-
Черно-подпалый — a
ta
tllWh-
Соболиный — A
y-llWh-
Афган
У афганской борзой встречается несколько окрасов. Это черный и черно-подпалый, что указывает на гены A
sи a
t. Распространен рыжий окрас различных оттенков. Многие такие особи имеют маску и соболиное затенение, что указывает на ген A
y. Получение черно-подпалых щенков от двух рыжих производителей (Литтл, 1957) также указывает на то, что именно доминантный ген А
уучаствует в образовании рыжего окраса. Голубой окрас у афганов встречается, но редко. Ссылаясь на эксперименты, проведенные в лаборатории Джексона, Литтл не исключает, что некоторые афганы обязаны своим рыжим окрасом гену е, а не A
y. Длинная шелковистая шерсть указывает на l.
Известны также осветленные вариации: желтые, палевые и почти белые. Это очень красивые и впечатляющие животные, особенно с черной маской. Литтл утверждает, что некоторые палевые вариации определяет ген c
chв комбинации с геном A
yили ее. В равной же степени здесь можно предположить и действие руфус-полигенов, которые ослабляют окрас до палевого или почти белого. Длинная шерсть усиливает эффект ослабления, т.к. более светлый подшерсток при длинной шерсти лучше виден и больше влияет на видимый окрас собаки, чем при короткой остевой шерсти, заслоняющей подшерсток. Некоторые осветленные особи, возможно, несут dd, особенно не имеющие маски. В комбинации cc
chили осветляющими руфус-полигенами такие особи будут полностью осветленными.
Окрас—
Генотип Черный — A
s-D-ll
Черно-подпалый — a
ta
tD-ll
Красный/рыжий — A
y-D-ll
Голубой — A
s-ddll
Кремовый — A
y-ddll
Эрдельтерьер
Эта порода представлена архитипичным чепрачным окрасом с хорошо выраженным V-образным чепраком и интенсивными подпалами на конечностях и морде. Встречается незначительная чалость на чепраке. Шерсть жесткая. Генотип: a
sa-Wh-.
Аляскинский маламут
Окрас в основном волче-серый, но варьирует от светло- до почти черно-серого. Некоторые особи имеют характерную светло-серую морду, другие имеют черную маску. Генотип: А- или a
sa- если эти два гена эквивалентны.
Американский кокер-спаниель
В основном, эта порода имеет такие же окрасы, что и английский кокер-спаниель. Поэтому, детально эти окрасы будут описаны ниже. Сомнительно, чтобы крупные генетические изменения могли произойти в процессе селекции американской версии. Значительно более обильное развитие длинных волос на конечностях американской породы указывает на влияние модификаторов. Оба кокера являются длинношерстными.
Американский фоксхаунд
Американский фоксхаунд значительно крупнее, чем английский, но генетически их окрасы идентичны. Литтл отмечает, что животные, лишенные белых отметин, становятся распространенными. Витни (1947) описал окрас типа мерль и назвал его «калико».
Окрас — Генотип
Рыжий или рыжевато-коричневый — A
y-S-
Чепрачный — a
sa-S-
Рыже-белый — A
y-s
p-
Чепрачный с белым — a
sa-s
p-
Американский стаффордшир-терьер
Эта порода несколько крупнее, чем стаффордшир-бультерьер. Генетика окрасов у них идентична.
Американский водяной спаниель
Не считая более мелких размеров порода идентична ирландскому водяному спаниелю и имеет генотип: A
s-bbkkll.
Анатолийская овчарка
Для породы наиболее характерны олений окрас и триколор. Последние характеризуется черной маской, рыжим туловищем и ирландской или пегой пятнистостью. Встречаются также серые или серо-коричневые по чепрачному типу собаки. Большинство особей оленьего окраса имеют черную маску, хотя выраженность ее вариабельна. Черные особи редки.
Окрас — Генотип
Черный — A
s-S-
Олений — A
y-S-
Серо-коричневый — a
sa-S-
Триколор — A
y-s
p-
Австралийский терьер
Собаки этой породы чепрачные, но, как и йоркширский терьер, имеют необычные цветовые вариации. Скорее всего они имеют одинаковую генетическую природу.
Австралийский шелковистый терьер
Порода также имеет только чепрачный окрас, еще более необычных серо-голубых и подпалых вариаций, сходных с окрасом йорка, что также указывает на одинаковую генетическую природу их окрасов.
Бассенджи
Окрас этой удивительной собаки обычно светло-рыжий, а вдоль спины проходит у некоторых черный ремень, что указывает на ген A
y. Возможно, что подпалые и черные собаки встречались раньше. Допускается ирландская белая пятнистость.
Окрас — Генотип
Черный — A
y-s
p-
Черно-подпалый — a
ta
ts-
Красный — A
y-s
i-
Бассет
Бассет может быть или рыжим (желтым, лимонный, красный) или черно-подпалым, даже ближе к чепрачному. При этом все собаки должны иметь отметины, характерные для пегости. Возможен крап, особенно в области морды, благодаря гену Т, экспрессия которого существенно ограничена.
Окрас — Генотип
Рыже-белый — A
y-s
p-
Триколор (черно-подпалый с белым) — a
sa-s
p-
Бигль
У бигля типичный чепрачный окрас, обусловленный геном a
sa. Здесь он относительно стабилен, хотя наблюдаются некоторые вариации степени развития чепрака и интенсивности пигментации как чепрака, так и поддала, что обусловлено полигонной вариабельностью. Неизменно присутствует белая пятнистость в виде «воротниковой» пятнистости или пегости. Таким образом генотип окраса бигля: a
sa-s
i- или a
sa-s
p-. Литтл (1957) отмечал, что некоторые индивидуумы имеют крап (ген Т).
В лаборатории Джексона описаны другие фенотипы. Так, бигли могли быть голубыми, желтыми, подпалы могли быть очень тусклыми. Это свидетельствует о работе генов b, d, e и, возможно, c
chсоответственно. Однако, перечисленные отклонения от обычного окраса встречались нечасто, примерно несколько особей на 100 щенков. Такая редкость означает нетипичность этих окрасов для породы в целом. Бигли — не единственная порода, где, при основном доминирующем окрасе, встречаются другие, более редкие цветовые вариации.
Бедлингтонтерьер
В соответствии с Литтлем (1957), общее осветление, типичное для данной породы, обязано доминантному гену G (раннее поседение). Голубые бедлингтоны рождаются черными и с возрастом светлеют. Допуская, что ген G присутствует во всех цветовых вариациях, истинную генетическую природу каждой из них можно определить только у щенка.
Обсуждая генетику пород, Литтл сделал ряд предложений. Так, он считает, что, хотя ген A
sотвечает за сплошной черный окрас, особи с осветлениями в местах подполов вторым геном в локусе имеют a
sa. Он также предположил, что ген c
chдает очень светлый голубой и очень светлый кремовый. Однако, поскольку c
chвесьма слабо влияет на черный пигмент, очень светло-голубой образуется скорее за счет гена d.
До конца еще не решено, что определяет рыжий (кремовый, палевый, песочный) окрас у бедлингтона: А
уили ее.
Все эти спорные вопросы могут быть разрешены, если это возможно, только при определении окраса у щенка до начала перецветания.
Окрас — Генотип
Голубой — A
s-B-D-G-
Голубо-подпалый — a
ta
tB-D-G-
Печеночный — A
s-bbD-G-
Коричнево-подпалый — a
ta
tbbD-G-
Песочный — A
y--D-G-
Кремовый — A
y--D-G-
Шерсть бедлингтона особенная. Она плотнил, необычной структуры. Нет никакой достаточной информации о характере наследования такой шерсти. Возможно, что это модификация жесткой шерсти (Wh), но это только предположение.
Бельгийская овчарка
По окрасу и по типу эта порода близка к немецкой овчарке. Порода подразделяется на три основные группы: грюнендаль, тервюрен и малинуа.
Бернская пастушья собака
Окрас бернской пастушьей собаки черно-подпалый с белым. В идеальном случае отметины располагаются на лбу, груди и лапах (типичная ирландская пятнистость). Шерсть длинная. Генотип: a
ta
tlls
i-.
Кунхаунд
Типичный генотип черно-подпалого окраса: a
ta
t.
Бладхаунд
Обычно это рыжие или черно-подпалые собаки (генетически скорее чепрачные). Некоторые рыжие собаки имеют маску. Редко встречаются коричнево-голубо-чепрачные.
Окрас — Генотип
Красный/рыжий — A
y--D-
Черно-подпалый (-чепрачный) — a
sa-B-D-
Коричнево-подпалый (-чепрачный) — a
sa-bbD-
Голубо-подпалый (-чепрачный) — a
sa-B-dd
Бордер-колли
Для этой породы характерно несколько окрасов, но, преимущественно, это черный с белыми отметинами. Генотип: A
s-S
i-.
Русская псовая борзая
Борзые бывают обычно муруго-пегими (соболе-пегими) с длинной шелковистой шерстью. Встречаются и черно-белые особи, но муругие предпочтительнее.
Окрас — Генотип
Черный — A
s-lls
p-
Муругий — A
y-lls
p-
Бостон-терьер
Основные окрасы породы — черный и тигровый с белыми отметинами и воротником. Симметричность пятен является важным породным признаком. Литтл (1957) считает, что такой окрас можно получить, отбирая собак, имеющих ирландскую пятнистость с максимальной экспрессией. В этом случае, ирландская пятнистость имеет тенденцию к симметричности, тогда как просто пегие собаки не имеют такой возможности. Литтл (1957) отмечает, что появление почти белых собак в этой породе получается в результате накопления гена s
w. Из этого следует, что некоторые типично окрашенные бостон-терьеры могли бы быть Ss
w. Некоторые черные особи являются результатом действия гена A
s, другие выглядят черными, хотя на самом деле — это просто сильно затемненные тигровые (E
br).
Окрас — Генотип
Черный — A
s--s
i-
Тигровый — A
y-E
br-s
i-
Боксер
Боксеры бывают только двух окрасов: рыжий (олений) и тигровый. Тигровый окрас очень вариабелен по количеству и интенсивности тигровин: от светло-тигровых, почти рыжих, до темно-тигровых, почти черных. У большинства собак этой породы есть маска. Некоторые имеют ирландскую пятнистость, но появляются и почти белые собаки, что обусловлено геном крайней степени пятнистости (s
w), который все еще встречается в этой породе, но нежелателен.
Окрас — Генотип
Тигровый — A
y-E
br-S-
Красный/рыжий/олений — A
y-E-S-
Тигровый с белым — A
y-E
br-s
i-
Рыжий с белым — A
y-Е-s
i-
Британский спаниель
Разрешенными для этой породы являются кофейно-белый и оранжево-белый окрасы. Осветленная или коричневая мочка носа всех оранжево-белых собак показывает, что эта порода полностью гомозиготна по гену b. Порядок расположения белых пятен характерен для генотипа s
ps
p. Белые отметины могут быть покрыты крапом. Литтл (1957) считает, что оранжевый окрас обусловлен геном е. Шерсть длинная.
Окрас — Генотип
Кофейный с белым — bbE-lls
p-tt
Кофейно-белый, с крапом — bbE-lls
p-Т-
Оранжево-белый — bbeells
p-tt
Оранжево-белый с крапом — bbeells
p-T-
Бультерьер
Среди бультерьеров встречаются и окрашенные и белые собаки. В настоящее время обсуждается вопрос, стоит ли скрещивать их между собой. Обычно, окрашенные особи черные, тигровые и рыжие (или темно-оленьи). Вариации последнего окраса обязаны руфус-полигенам. У красных и оленьих собак может быть в разной степени выраженная маска. Многие животные имеют ирландскую пятнистость я, возможно, несут s
is
i. Бриггс и Калисс (1942) отмечают, что черно-подпалые и голубые собаки появились в породе позже.
Белые вариации обязаны генотипу s
ws
w. Об этом говорят пигментированные пятна на голове у некоторых белых животных (Бриггс, 1940; Литтл, 1957).
Все гены окраса, присущие породе бультерьер, конечно, присутствуют в геноме белого буля, но их действие подавлено геном крайней степени белой пятнистости. Наличие у некоторых белых особей крупных пятен позволяет определить у них некоторые гены.
Окрас — Генотип
Черный — A
s-Е-s
i-
Олений/рыжий — A
y-Е-s
i-
Тигровый — A
y-E
br-s
i-
Бульдог
Обычно это рыжие, соболиные или тигровые собаки с маской или без нее. Возможно, что желтовато-коричневый окрас является результатом действия или гена шиншилла c
ch, который ослабляет интенсивность желтого пигмента, или ослабляющих руфус-полигенов. Следует отметить наличие белой пятнистости всех вариантов и степеней, что позволяет предположить присутствие всех трех аллелей белой пятнистости: s
i, s
pи s
w. Белые участки могут быть с крапом (ген Т).
Окрас — Генотип
Красный — A
y-E-S-
Соболиный — A
y-E-S-
Тигровый — A
y-E
br-S-
Красно-белый — A
y-E-s
p-
Соболино-белый — A
y-Е-s
p-
Тигровый с белым — A
y-E
br-s
p-
Бульмастиф
Генетика окрасов в этой породе идентична с таковой у боксеров». Но у бульмастифа не бывает белых отметин. Интенсивность желтой пигментации варьирует от красной до желтовато-коричневой под влиянием руфус-полигенов. Черная маска обязательна и хорошо выражена.
Окрас — Генотип
Тигровый — A
y-E
br-
Красный/желтовато-коричневый — A
y-Е-
Кернтерьер
Еще одна трудная для генетического анализа порода. Литтл (1957) считает, что изначально, генетически, окрас кернтерьера тигровый, но модификация его такова, что полосы все ломаются и путаются. Эффект усугубляется густой лохматой шерстью. Фенотипически, окрас варьирует от почти черного до почти рыжего со множеством промежуточных вариантов в соответствии с соотношением черных и желтых полос. Интенсивность желтого фона тоже варьирует от песочного до почти белого, через кремовый в результате действия руфус-полигенов. Контраст между темной головой и туловищем обусловлен наличием черной маски, а также более длинными волосами на туловище, чем на голове.
Чисто красные, рыжие, песочные; палевые собаки имеют ген A
y, экспрессию которого модифицируют руфус-полигены. Возможно, что некоторые вариации тигрового, рассмотренный выше, на самом деле являются соболиными.
Окрас — Генотип
Тигровый — A
y-E
br-ll
Красный/песочный — A-E-ll
Кавалер-кинг-чарлз-спанлель
Окрасы в этой породе четко определены: черно-подпалый и красный, который обусловлен действием как A
y, так и ее. Их можно легко различить. Если при скрещивании двух красных собак получаются только красные щенки, то родители несут ее, если же получаются и черно-подпалые, то, по крайней мере, один из производителей несет A
y.
Как черно-подпалый, так и красный окрасы могут быть комбинированы с белой пятнистостью. Такие окрасы называются триколор и бленхейм соответственно.
Окрас — Генотип
Черно-подпалый — a
ta
tE-llS-
Триколор — a
ta
tE-lls
p-
Красный/рубиновый — -eellS-
Бленхейм — -eells
p-
Чесапик-бей-ретривер
Собаки этой породы полностью коричневого окраса различных оттенков, от темно-коричневого до цвета «жухлой травы». Основной генотип: A
s-bb. Литтл (1957) предположил, что осветленные особи имеют ген шиншилла c
ch. В таком случае, исходя из влияния его на коричневый окрас у других животных, для ретриверов с геном c
chследует ожидать «хаки»-подобного окраса.
Шерсть имеет тенденцию виться, но нет достаточных сведений о наследуемости этого признака. Степень волнистости можно контролировать путем отбора.
Чихуахуа
Для этих изящных маленьких собачек характерен широкий спектр окрасов: черный, черно-подпалый, соболиный, коричневый, коричнево-подпалый и коричнево-соболиный. Литтл (1957) отметил, что черные щенки могут быть получены от двух рыжих собак. Следовательно, у чихуахуа, наряду с геном A
y, рыжие окрасы могут быть образованы в результате экспрессии гена e. Однако, неизвестно, насколько этот ген распространен в природе. Рыжие окрасы настолько сильно варьируют по интенсивности, что можно предположить наличие в генофонде породы гена c
ch- или/и многочисленных ослабляющих руфус-полигенов, которые так уменьшают количество желтого пигмента, Что собаки становится почти белой. Голубые особи известны, но очень редки. Распространена ирландская пятнистость и пегость. Литтл отмечал, что могут встречаться и крайние степени пятнистости, вплоть до полностью белых собак. Все перечисленные окрасы встречаются как среди короткошерстных, так и среди длинношерстных вариаций.
Перечисленные в списке генотипы окрасов можно скомбинировать с белой пятнистостью путем замены S на s
i, s
pили s
wи/или с длинношерстностью, соответственно заменяя L на l.
Окрас — Генотип
Черный — A
s-B-D-L-S-
Черно-подпалый — a
ta
tB-D-L-S-
Кофейный — A
s-bbD-L-S-
Кофейно-подпалый — a
ta
tbbD-L-S-
Голубой — As-B-ddL-S-
Голубо-подпалый — a
ta
tB-ddL-S-
Соболиный — A
y-B-D-L-S-
Коричнево-соболиный — A
y-bbD-L-S-
Красный (черная мочка носа) — A
y-B-D-L-S-
Красный (коричневая мочка носа) — A
y-bbD-L-S-
Китайская хохлатая собачка
Типичный пример безволосой собаки. Некоторое количество волос встречается на голове, нижней части конечностей и на хвосте. Напротив, мексиканские голые собаки фактически лишены шерсти. Судя по длине оставшихся волос, предки китайской собачки были длинношерстными. Туловище может быть одноцветным или пятнистым. В последнем случае, кожа имеет темную пигментацию о депигментированными пежинами, часто покрытыми крапом. При отсутствии волос у собак этой породы трудно рассуждать о генотипах окрасов.
Окрас — Генотип
Однотонный — Hr-llS-
Пятнистый — Hr-lls
p-T
Чау-чау
Для чау-чау характерны черный, голубой и различные оттенки рыжего окраса. Черный образуется за счет A
s, голубой — b. В соответствии с исследованиями Литтля, красные собаки могут иметь как A
y, так и ее. Олений, кремовый, палевый окрасы могут получаться в результате действия гена d или ослабляющих руфус-полигенов. Шерсть у чау длинная и очень хорошо развита по всему корпусу.
Окрас — Генотип
Черный — A
s-D-ll
Голубой — A
s-ddll
Красный — A
y-D-ll
Кремовый — A
y-ddll
Кламбер-спаниель
У этого спаниеля так много белого, что можно предположить скорее наличие s
w, чем s
p. Рыжие или желтые участки различных оттенков чаще всего расположены на голове. Литтл (1957) считает, что желтый пигмент пятен образуется за счет гена е. Мочка носа обычно кофейная или телесного цвета, что свидетельствует о наличии гена b. Т.е. генотип окраса кламбер-спаниеля: bbeells
ws
w.
Жесткошерстный грифон
Кофейная или каштановая собака с полностью прокрашенной головой и чалым корпусом. Шерсть длинная и грубая. Возможно, что генотип: A
s-bblls
p-T-Wh- пли As-bb-lls
ws
wT-Wh-. Эффект чалости усиливается длинной жесткой шерстью.
Курчавый ретривер
Собаки бывают как черные, так и кофейные. Генотипы этих окрасов A
s-В- и A
s-bb соответственно. Основной характерной особенностью породы является присутствие коротких плотных завитков по всему телу. Возможно, что этот признак обусловлен тем же геном, что и у ирландского водяного спаниеля. Тогда генотип можно описать следующим образом: черный — A
s-B-kk и кофейный — A
s-bbkk. Однако, Бернс и Фразер (1966) обнаружили, что в результате скрещивания курчавого ретривера с дратхааром, щенки имели курчавую шерсть. Следовательно, курчавость ретривера определяется доминантным геном, возможно, не связанным с геном k.
Такса
У такс обнаруживается огромное количество вариантов окраса и типов шерсти. Большинство такс рыжие, вероятнее всего за счет A
y, т.к. имеют маску и черное затенение на спине. Распространены также черно-подпалые и кофейно-подпалые собаки. Гораздо более редкими считаются мраморные собаки (гетерозиготные по М:Mm).
Необычным, но допустимым окрасом является волче-серый или серый с проседью (кабаний), который определяется или геном агути А или геном чепрачности a
sa, экспрессия которого ослаблена действием модификаторов.
Для такс характерны три типа шерсти: гладкая, длинная и жесткая. Два последних определяются генами l и Wh соответственно.
Представляем наиболее полный список окрасов и их генотипов.
Окрас — Генотип
Черно-подпалый — a
ta
tB-L-mmwhwh
Кофейно-подпалый — a
ta
tbbL-mmwhwh
Рыжий/соболиный (черная мочка носа) — A
y-B-L-mmwhwh
Рыжий/соболиный (кофейная мочка носа) — A
y-bbL-mmwhwh
Мраморный (черно-голубой) — a
ta
tB-L-Mmwhwh
Мраморный (красно-желтый) — A
y--L-Mmwhwh
Красный, длинная шерсть — A
y-B-llmmwhwh
Мраморный длинная шерсть — a
ta
tB-llMmwhwh
Черно-подпалый, длинная шерсть — a
ta
tB-llmmwhwh
Кофейно-подпалый, длинная шерсть — a
ta
tbbllmmwhwh
Черно-подпалый, жесткая шерсть — a
ta
tB-L-mmWh-
Кофейно-подпалый, жесткая шерсть — a
ta
tbbL-mmWh-
Красный, жесткая шерсть — A
y--L-mmWh-
Мраморный, жесткая шерсть — a
ta
tB-L-MmWh-
Далматин
Уникальный окрас далматинов получен в результате бесчисленного множества селективных скрещиваний. Литтл (1957) показал, что генотип далматина s
ws
wT-. Собаки этой породы крапчатые за счет гена Т, но экспрессия его модифицирована в результате селекции таким образом, что обычный крап превратился в равномерно разбросанные по всему телу пигментированные круглые пятна. Обычно, эти пятна черного или кофейного цвета. Известны также желтые вариации, благодаря гену e, по мнению Литтля. Особи с желтым крапом могут иметь как черную, так и коричневую мочку носа.
Окрас — Генотип
Черный — A
s-B-E-s
ws
wT-
Кофейный — A
s-bbE-s
ws
wT-
Желтый (черная мочка носа) — -B-ees
ws
wT-
Желтый (коричневая мочка носа) — -bbees
ws
wT-
Денди-динмонт-терьер
Окрасы, присущие данной породе, трудно анализировать, благодаря цветовым изменениям, которые претерпевают собаки от рождения до созревания. В соответствии с Литтлем (1957) горчичные собаки рождаются темно-соболиными, а перечные — черно-подпалыми. В обоих случаях взрослые особи значительно светлее, чем щенки, что характерно для экспрессии гена раннего поседения G. Литтл отметил, что некоторые перечные вариации несут не a
t(черно-подпалый), а A
s. В соответствии с Литтлем (1957), генотип 2-х вариаций окраса может быть таков: A
s-G-ll A
y-G-ll для горчичного и a
ta
tG-ll для перечного.
Дирхаунд
Литтл (1957) считает, что окрас этих собак изначально тигровый, образованный не как обычно, с четким разграничением черных и рыжих полос, а путем смешения черных и желтых волос. Удлиненная, спутанная грубая, вьющаяся шерсть усиливает эффект. Литтл утверждает, что полосы все-таки можно обнаружить у новорожденных щенков или стриженых собак. Цветовые вариации получаются в результате различных соотношений темных я светлых волос и интенсивности пигментации последних. Возможно, что генотипы этих собак A
y-E
br-Wh- и a
sa-E
br-Wh-. Сплошные желтые, рыжие, песочные и палевые особи (обычно с черной маской) несут генотип A
y-E-Wh-. Неясно пока, какой ген отвечает за длину шерсти L или l.
Доберман
Доберман является архитипичной подпалой собакой. Различают три цветовые вариации: черно-подпалый, кофейно-подпалый и голубо-подпалый. Подпалы должны быть яркими, четкими и в строго определенных местах. Лиловый окрас, или изабелла, может возникать время от времени, так как в породе есть и ген b, и ген d. Лиловый окрас получается, если в геноме присутствуют оба эти гена в гомозиготном состоянии. Иногда изабеллу путают с голубым, иногда появление его вообще обескураживает.
Окрас — Генотип
Черный — a
ta
tB-D-
Коричневый — a
ta
tbbD-
Голубой — a
ta
tB-dd
Изабелла — a
ta
tbbdd
Дункер
Эта необычная, интересная порода норвежского происхождения заслуживает особого внимания, так как собаки этой породы гетерозиготны по гену мерль М. Окрас дункера голубовато-серый с неровными рваными черными пятнами, равномерно разбросанными по корпусу. Встречаются собаки с белыми отметинами по типу ирландской пятнистости. Генотип: A
s-Mms
i-.
Элкхаунд
Окрас этой очень крепкой собаки близок к волче-серому. Основой окраса является чепрак, но значительно более осветленный, чем стандартный, за счет большого количества желтых волос различных оттенков у разных особей, которые перемежаются с черными на чепраке. Литтл предположил, что серый элкхаунд имеет ген дикого типа A, однако автор настоящей книги склонен заменить его геном чепрачности a
sa(см. предыдущую главу). Присутствует черная маска. Известны черные собаки, но они редки. Литтл описал кофейный окрас (a
sa-bb).
Английский кокер-спаниель
Для английского кокера допустим широкий спектр окрасов. Фактически, почти все известные окрасы, сопряженные с пегостью и, в ряде случаев, с крапчатостью встречаются у этих собак. Крапа обычно не много, хотя некоторые собаки, в результате гиперразвития тиковости, становятся чалыми. Обычными считаются черно-подпалые, черные, кофейные, рыжие. Последние как за счет A
y, так и е. В породе присутствуют оба этих гена. В соответствии с Литтлем (1957), основная часть рыжих собак имеет е, но некоторая часть имеет A
y, так как иногда от двух красных кокеров получаются черные щенки. Литтл описал рождение голубых, что говорит о циркуляции в породе гена d в очень незначительных пропорциях.
Интенсивность окраса рыжих собак и подпалов у черно-подпалых варьирует от красного до палевого в результате действия руфус-полигенов. Нельзя исключить также наличие гена c
ch(шиншилла). У некоторых почти белых животных, окрашенные пятна присутствуют только на ушах, что свидетельствует о наличии в генофонде породы аллеля s
w, наряду с s
p. Такая пятнистость возникла в результате или крайней степени экспрессии сочетания s
ps
pпод действием модификаторов или работы гена s
wв сочетании s
ps
wили s
ws
w.
Шерсть у английских кокеров длинная, что свидетельствует о наличии гена Е. Многие спаниели имеют я ратной степени выраженную волнистость шерсти. Кик правило шерсть больше волнится на голове и конечностях. Витни утверждает (1947), что у кокера прямая шерсть доминантна по отношению к волнистой, но допускает, что широкий спектр вариабельности волнистости создает проблемы в диагностике волнистых индивидуумов.
Окрас — Генотип
Черный — A
s-B-E-llS--
Черно-белый — A
s-B-E-lls
p-tt
Черно-белый, крапчатый — A
s-B-E-lls
p-T-
Черно-подпалый — a
ta
tB-Е-llS--
Триколор — a
ta
tB-E-lls
p-tt
Триколор с крапом — a
ta
tB-E-lls
p-T-
Кофейный — A
s-bbE-llS--
Кофейно-белый — A
s-bbE-lls
p-tt
Кофейно-белый с крапом — A
s-bbE-lls
p-T-
Кофейно-подпалый — a
ta
tbbE-llS--
То же с белым — a
ta
t-bbE-lls
p--
То же с белым, крапчатый — a
ta
tbbE-lls
pT-
Красный (черная мочка носа) — -B-eellS--
То же с белым — -B-eells
p-tt
То же с белым, с крапом — -B-eells
p-T-
Красный (коричневая мочка носа) — -bbeellS--
То же с белым — -bbeells
p-tt
То же с белым, с крапом — -bbeells
p-Т-
Английский сеттер
В этой породе встречается несколько окрасов. Черный, кофейный и лимонный составляют основу окраса, на которую накладываются белая пятнистость и крапчатость, настолько выраженные, что собаки кажутся чалыми. Степень выраженности пятнистости варьирует от пегого до почти белого окраса. В последнем случае на белых участках так много крана, что делает собаку полностью чалой. Предполагается, что в породе присутствует как s
p, так и s
w. Черно-подпалый с белым известен как триколор. Литтл (1957) доказал, что лимонный окрас обязан гену е. Шерсть длинная (l).
Окрас — Генотип
Черно-крапчатый — A
s-В-Е-lls
p-Т-
Кофейно-крапчатый — A
s-bbE-lls
p-T-
Черный триколор — a
ta
tB-E-lls
p-T
Кофейный триколор — a
ta
tbbE-lls
p-Т-
Лимонный (черная мочка носа) — -B-eells
p-T-
Лимонный (коричневая мочка носа) — -bbeells
p-T-
Английский спрингер-спаниель
Как и у английского кокер-спаниеля, у спрингер-спаниеля существует множество различных окрасов. Это черный и кофейный, как сплошной, так и подпалый; красный и желтый в соответствии с количеством руфус-полигенов; крапчатый и нет: пятнистый, пегий и пр.
Окрас — Генотип
Черный — A
s-B-E-llS-
Черно-белый — A
s-B-E-lls
p-tt
Черно-белый с крапом — A
s-B-E-lls
p-T-
Черно-подпалый — a
ta
tbbE-llS-
То же с белым — a
ta
tbbE-lls
p-tt
То же с белым с крапом — a
ta
tbbE-lls
p-T-
Кофейный — A
s-bbE-llS--
Кофейно-белый — A
s-bbE-lls
p-tt
То же с крапом — A
s-bbE-lls
p-T-
Кофейно-подпалый — a
ta
tbbE-llS--
То же — a
ta
tbbE-lls
p-tt
То же с белым, с крапом — a
ta
tbbE-lls
p-T-
Красный (черная мочка носа) — -B-eellS--
То же с белым — -B-eells
p-tt
То же с белым, с крапом — -B-eells
p-T-
Красный (коричневая мочка) — -bbeellS--
То же с белым — -bbeells
p-tt
То же с белым, с крапом — -bbeells
p-T-
Английский той-терьер
Это миниатюрный манчестерский терьер с генотипом a
ta
t.
Филд-спаниель
Как и у других спаниелей, в этой порода множество окрасов. Предпочтение отдается сплошным окрасам без белых пятен, хотя пятнистые собаки не запрещены. Список окрасов и их генотипов такой же, что у английского кокер-спаниеля.
Финский шпиц
Окрас желто-рыжий, соболиный с симметричным затенением. Шерсть длинная. Генотип: A
y-ll.
Прямошерстный ретривер
Для породы характерны только дна окраса. Это черный и кофейный. Их генотипы: A
s-B- и A
s-bb соответственно.
Фоксхаунд
Типичная чепрачная пегая собака. Встречаются, однако, и черные, рыжие, светло-рыжие животные (обязательно пегие).
Окрас — Генотип
Черный — A
s-s
p-
Рыжий — A
y-s
p-
Чепрачный — a
saa
sas
p-
Французский бульдог
При оленьем окрасе французский бульдог имеет черную маску, следовательно, несет ген А
у. Очень темная тигровость бульдога есть результат действия гена E
br. Фактически, большинство черных животных этой породы являются на самом деле тигровыми с минимальным количеством желтых пестрин или без них. Характерна белая пятнистость, которая варьирует от ирландской до почти белого окраса. Поэтому, можно допустить наличие в породе всех трех аллелей пятнистости (s, s
pи s
w).
Окрас — Генотип
Олений — A
y-E-S-
Тигровый — A
y-E
br-S-
Олений с белым — A
y-Е-s
p-
Тигровый с белым — A
y-E
br-s
p-
Немецкая овчарка
Немецкие овчарки бывают различных окрасов. И в каждом из них наблюдаются значительные вариации. Так, соболиный может варьировать от рыжего с незначительным вкраплением черных волос до стильно затененного. Чепрачный окрас варьирует от совсем светлого с большим количеством рыжих волос на чепраке, перемежающихся с черными, до почти черного окраса с почти незаметными рыжими подпалами. Черно-подпалый окрас значительно менее вариабелен и варьирует в сторону уменьшения площади подпалов. Во всех случаях маска обязательна. Интенсивность желтой пигментации варьирует от золотого до светло-палевого, почти белого.
На основании вышесказанного следует, что порода имеет все аллели локуса агути (А
у, А
у, a
saи a
t), экспрессия которых варьирует в зависимости от модифицирующих генов. Особенно выражены вариации в экспрессии a
sa, тогда как вариабельность проявления гена A
yтакая же, как у других пород собак. У чепрачных немецких овчарок чепрак может быть узким или широким, сплошным черным или с примесью различного количества желтых волос. Чепрак является основной характерной чертой окраса. Кроме того чепрачный окрас характеризуется также темными отметинами на груди, голове, плечах и вдоль бедер.
Таким образом, чепрачность проявляется в огромном числе модификаций, тогда как рыжие окрасы более стабильны: степень интенсивности желтой пигментации варьирует от золотистого до светло-палевого.
Среди немецких овчарок встречаются особи с волче-серым окрасом. Генетическую природу его можно интерпретировать двояко. Во-первых, это может быть модификация чепрачного окраса (a
sa), когда черные волосы чепрака интенсивно перемешаны с желтыми, что в целом делает собаку волче-серой. Во-вторых, нельзя исключить наличие в породе гена дикого типа A (агути). К сожалению, Литтл считает, что гены a
saи a
tявляются одним геном, а аллель дикого типа A он называет a
wи считает рецессивным.
Светло-палевые или белые с темными отметинами, которые время от времени появляются среди немецких овчарок скорее всего обязаны своим окрасом гену A
yв комбинации с геном c
ch(как предполагает Литтл) или с осветляющими руфус-полигенами. Темные отметины в виде маски и ремня на спине не подвержены влиянию ни c
ch, ни полигонов.
Необходимо добавить, что Виллис (1976) опубликовал результаты селекционной работы, где он доказывает, что у немецкой овчарки черный окрас наследуется рецессивно. От двух черно-подпалых особей получались черные щенки. Енсен считает, что этот окрас обязан рецессивному гену а из серии агути, аналогично гену черного окраса а многих млекопитающих.
Окрас — Генотип
Соболиный — A
y-L-
Чепрачный — a
sa-L-
Черно-подпалый — a
ta
tL-
Соболиный длинношерстный — A
y-ll
Чепрачный длинношерстный — a
sa-ll
Черно-подпалый длинношерстный — a
ta
tll
Курцхаар
Эти собаки имеют или сплошной кофейный окрас или кофейно-пегий. У многих животных на белом фоне произвольно разбросаны мелкие кофейные пятна.
Окрас — Генотип
Кофейный — A
s-bbS--
Кофейно-белый — A
s-bbs
p-tt
Кофейно-белый с крапом — A
s-bbs
p-T-
Дратхаар
Окрасы полностью совпадают с окрасами курцхаара. Различия состоят в типе шерсти. Жесткая шерсть, в данном случае, обусловлена геном Wh. Такая шерсть делает собаку с крапом чалой (ген Т). Некоторые исследователи полагают, что наряду с геном тиковости Т должен существовать отдельный ген чалости, но очевидных экспериментальных доказательств пока нет.
Золотистый ретривер
Породе присущ только рыжий окрас во всех его проявлениях: от насыщенного золотого до светло-палевого. Генетически он определяется геном е, следовательно, основной генотип, характеризующий породу, будет eell. Вариации интенсивности окраса зависят от руфус-полигенов. Однако, некоторые животные настолько осветлены, что напрашивается вывод об участии гена c
ch.
Сеттер-гордон
Исключительно черно-подпалые собаки. Шерсть у них средней длины. Возможно, что действие ll модифицировано таким образом, что получилась недлинная прилегающая шерсть. Окрас подпалов обычно красный. Литтл (1957) описал собаку с осветленными подпалами. Он приписал этот эффект действию гена c
ch, так как такие животные дискретно выщепляются при отсутствии промежуточных вариантов, Однако, не стоит исключать и наличия руфус-полигенов. И Литтл и Винг утверждают, что рыжий окрас в этой породе отсутствует. Если бы такие особи возникли, их генотип был бы a
ta
teell. Генотип сеттера-гордона a
ta
tEEll.
Немецкий дог
Стандартные окрасы: олений, который варьирует от рыжего до светло-палевого, тигровый, черный и голубой. Олений окрас обязан гену A
y, который, взаимодействуя с E
br, дает тигровый, осветленный за счет большой площади рыжего. Голубой — это ослабленный черный. Допускаются голубо-тигровые собаки, но они редки из-за непривлекательного внешнего вида. Во всех случаях маска обязательна, только у осветленных рыжих или голубых маска тоже осветлена.
Встречаются мраморные окрасы (арлекин), когда на белом фоне разбросаны черные разорванные пятна по всей голове и корпусу. В соответствии с Литтлем, окрас получается в результате комбинации одного из генов пятнистости, возможно s
p, и гена мерль (М) в гетерозиготном состоянии. Обычные пегие собаки (mms
p-) должны были бы иметь на белом фоне пигментированные пятна, но недостаточно разорванные и более ровные по краю. Голубой арлекин вместо черных пятен имеет голубые.
Похоже, что мраморный окрас дога отличается от соответствующего у такс и колли. Черные пятна у мраморных такс и колли расположены не на белом, а на голубом фоне. Возможно, что окрас арлекин у дога образован геном H
h, описанный Шайбмен и Брюнбау (1976) и названный мраморный арлекин. У индивидуумов H
hh основной фон белый или почти белый.
Окрас — Генотип
Черный — A
s-D-E-mmS-
Олений (черная маска) — A
y-D-E-mmS-
Тигровый — A
y-D-E
br-mmS-
Голубой — A
s-ddE-mmS-
Ослабленный олений — A
y-ddE-mmS-
Голубо-тигровый — A
y-ddE
br-mmS-
Черный арлекин — A
s-D-E-Mms
p-
Голубой арлекин — A
s-ddE-Mms
p-
Грейхаунд
Грейхаунд имеет множество окрасов. Все ординарные, распространенные окрасы собак представлены в этой породе, за исключением подпалых и кофейных. Поэтому, не стоит особо подробно обсуждать окрасы грейхаундов. Следует только отметить, что рыжий окрас варьирует у них от насыщенного рыжего до палевого через олений. Эти вариации обусловлены как действием гена c
ch, так и руфус-группы. Наблюдая большое разнообразие пятнистых грейхаундов, Литтл пришел к выводу, что в породе присутствуют все три аллеля белой пятнистости. Наиболее распространенным из них он считает s
p.
Окрас — Генотип
Черный — A
s-D-E-S-
Голубой — A
s-ddE-S-
Черно-тигровый — A
y-D-E
br-S-
Голубо-тигровый — A
y-ddE
br-S-
Красный/ желтый — A
y-D-E-S-
Олений/палевый — A
y-ddE-S-
Черно-пегий — A
s-D-E- s
p-
Голубо-пегий — As-ddE- s
p-
Черно-тигровый с белым — Ay-D-E
br-s
p-
Голубо-тигровый с белым — Ay-ddE
br-s
p-
Красный/желтый с белым — Ay-D-E-s
p-
Олений/палевый с белым — Ay-ddE-s
p-
Брюссельский грифон
Стандартными считаются черные, черно-подпалые и рыжие с маской. Шерсть прямая или жесткая. Собаки с жесткой шерстью выглядят скорее как длинношерстные.
Окрас — Генотип
Черный — A
s-whwh
Черно-подпалый — a
ta
twhwh
Красный — A
y-whwh
Черный, жесткая шерсть — A
s-Wh-
Черно-подпалый, жесткая шерсть — a
ta
tWh-
Красный, жесткая шерсть — A
y-Wh-
Халденстрёваре
Ладно скроенная, среднего размера чепрачно-пегая собака. Генотип: a
sa-s
p-.
Венгерская выжла
Рыжие собаки различных оттенков, от насыщенного песочно-желтого до медно-золотого. Мочка носа и губы коричневые. Генотип: A
y-bb.
Харрьер
Уменьшенная копия фоксхаунда.
Испанский паденго
Рыжая или желто-пегая собака. Мочка носа коричневая благодаря bb. Шерсть обычно короткая, но встречается и длинношерстные экземпляры, и жесткошерстные.
Окрас — Генотип
Красный/рыжий — A
y-L-s
p-whwh
Красный/рыжий, длинношерстный — A
y-lls
p-whwh
Красный/рыжий, жесткошерстный — A
y-L-s
p-Wh-
Ирландский сеттер
У ирландского сеттера бывает только рыжий окрас. Литтл (1957) показал, что полное отсутствие черного пигмента в волосах говорит о наличии только гена е. Хотя шерсть и не достаточно обильная и не вполне длинная, многие свойства ее позволяют считать, что структуру шерсти определяет ген l.
Если это так, то генотип породы: A
s-eell.
Максимальная концентрация усиливающих руфус-полигенов обеспечивает очень насыщенный красный окрас. Винг (1950) отмечал, что о прошлом от рыжих родителей получались черные щенки. Литтл привел такие статистические данные: на 1197 щенков приходится один черный. По свидетельствам собаководов могут получаться также черно-подпалые, черные и соболиные щенки.
Это свидетельствует о том, что некоторая незначительная часть поголовья имеет не A
s-ee, а A
y-ЕЕ.
Ирландский терьер
Ирландский терьер имеет рыжий окрас и жесткую шерсть. Интенсивность желтого варьирует от красного до желто-рыжего, что определяется руфус-полигенами. Генотип: A
y-WhWh.
Ирландский водяной спаниель
Кофейный окрас и длинная вьющаяся шерсть — единственное, что допустимо в этой породе. Витни (1957) опубликовал данные, которые доказывают, что ген курчавости (терминология Витни) наследуется как рецессивный. Генотип: A
s-bbkkll.
Ирландский волкодав
Литтл (1957) считает, что у ирландского волкодава, как и у дирхаунда, тигровый окрас нечетко проявляется из-за жесткой грубой шерсти и изломанных тигровин. Однако, утверждать, что при таком окрасе генотип должен быть a
sa-E
br- преждевременно.
Стандартными являются также черный, рыжий, олений и белый.
Литтл считает, что белые особи на самом деле светло-палевые в результате взаимодействия гена A
yс c
chили руфус-полигенами. До сих пор неясно, присутствует ли ген l в породе.
Окрас — Генотип
Черный — A
s-Е-
Тигровый — A
y-E
br-
Красный/олений — A
y-Е-
Левретка
Здесь допустимы все окрасы грейхаунда за исключением тигровых. Это означает, что ген E
brисключен из генофонда данной породы.
Японский хин
Японские хины бывают черно-пегими и рыже-пегими. Рыжие особи могут быть как полностью свободны от черного пигмента, так и затенены в различной степени (соболиный тип). Шерсть длинная, шелковистая.
Окрас — Генотип
Черно-белый — A
s-lls
p-
Рыже-белый — A
y-lls
p-
Кеесхонд
По стандарту собаки должны быть волче-серыми или пепельно-серыми (А). Возможно, что такой окрас образуется за счет взаимодействия генов чепрачности (a
sa) и осветляющих руфус-полигенов, делающих подпалы палевыми, а чепрак волче-серым. Шерсть длинная. Генотип: A-ll или a
sa-ll.
Керри-блю-терьер
Щенки рождаются черными и постепенно становятся голубыми. Голубой окрас взрослых варьирует от темно-серого до серебристого. Такая возрастная трансформация окраса является характерной для гена G, описанного Литтлем. У некоторых, но не у всех, имеется маска. Возможно, что ген G не может осветлять слишком интенсивную пигментацию маски. Шерсть длинная, плотная, вьющаяся. Генотип: A
s-G-ll.
Норфолк-терьер
Обычно это Ay-рыжие собаки с легким красноватым оттенком. Черное затенение может возникать на голове, хвосте и боках. Из-за этого такой окрас называется «гризли». Встречаются также черно-подпалые собаки. Шерсть на вид жесткая.
Окрас — Генотип
Красный — A
y-Wh-
Гризли — A
y-Wh-
Черно-подпалый — a
ta
tWh-
Норвежский бухунд
Окрасы в этой породе могут быть волче-серыми, светло-рыжими до бледно-желтого и черными. У некоторых животных имеется маска.
Окрас — Генотип
Черный — A
s-
Рыжий — A
y-
Волче-серый — A-
Норвич-терьер
Окрасы и качество шерсти этого небольшого терьера как и у норфолк-терьера (см. выше).
Староанглийская овчарка (бобтейл)
Это еще одна порода, которая, по мнению Литтла, имеет ген раннего поседения (G). Литтл признает, однако, что некоторые особи могут быть просто голубыми за счет ослабляющего гена d. Он не исключает также присутствие гена мерль (М). Но вопрос этот спорный. Большая часть староанглийских овчарок имеет белые отметины по типу «воротниковой» пятнистости, хотя по стандарту допускаются и сплошные окрасы. Шерсть длинная и косматая, что затрудняет четкую идентификацию окраса. Генотип типичной собаки можно представить как: A
s-G-lls
p-.
Папильон
Это одна из наиболее изящных собачек. Папильоны всегда белые с черными или рыжими пятнами или черно-подпалые с белым.
Окрас — Генотип
Черный (с белым) — A
s-lls
p-
Красный (с белым) — A
y-lls
p-
Черно-подпалый (с белым) — a
ta
tlls
p-
Пекинес
Пекинес — порода, которая имеет много различных окрасов. Наиболее популярны красные и соболиные собаки. У многих животных имеется черная маска. Встречаются светло-палевые и почти белые особи. Такие окрасы определяются или генотипом A
y-c
chили большим количеством ослабляющих руфус-полигенов. В соответствии с Литтлем (1957), при скрещивании двух рыжих собак можно получить черное потомство. Следовательно, наряду с геном A
yв породе должен присутствовать ген е. Степень пятнистости у пекинесов сильно варьирует, что говорит о наличии в породе всех трех генов белой пятнистости (s, s
p, s
w). Литтл описал голубого пекинеса (ген d). Время от времени появляются почти или полностью белые собаки. Шерсть длинная, ниспадающая.
Окрас — Генотип
Черный — A
s-llS-
Красный — A
y-llS-
Соболиный — A
y-llS-
Черно-белый — A
s-lls
p-
Рыже-белый — A
y-lls
p-
Соболе-белый — A
y-lls
p-
Кинг-чарльз-спаниель
Эта порода, по существу, является уменьшенной копией кавалер кинг-чарльз-спаниеля (см. выше).
Лабрадор-ретривер
Стандартом приняты черные и рыжие собаки, хотя встречаются кофейные особи. Почти наверняка, рыжие окрасы Лабрадора обязаны гену е (Бернс и Фразер, 1966). Вариации интенсивности желтой пигментации происходят за счет руфус-полигенов.
Окрас — Генотип
Черный — A
s-В-ee
Кофейный — A
s-bbee
Рыжий (черная мочка носа) -B-ee
Рыжий (коричневая мочка носа) -bbee
Лейкленд-терьер
Порода близка по происхождению к эрдельтерьеру, но имеет более широкий спектр окрасов. Это черный, голубой, кофейный, черно- и голубо-чепрачный, рыжий, пшеничный и красный гризли. Генотип красного гризли спорен. На самом деле этот окрас может относиться или к светло-чепрачному или к соболиному. Шерсть жесткая.
Окрас — Генотип
Черный — A
s-D-Wh-
Чепрачный — a
sa-D-Wh-
Голубой — A
s-ddWh-
Голубо-чепрачный — a
sa-ddWh-
Красный — A
y-D-Wh-
Большой мюнстерлендер
Белая, среднего размера собака с редкими черными пятнами и крапом. Крап может варьироваться от нескольких пятен до малости. Шерсть длинная. Генотип: A
s-lls
p-T-.
Мальтийская болонка
Предпочтителен чисто белый окрас, хотя допустимы желтоватые отметины. Литтл (1957) предположил, что белый окрас болонки обусловлен крайней степенью проявления аллеля белой пятнистости s
wи/или осветляющими руфус-полигенами, скорее, чем комбинацией A
yили е c
chc
ch. Он утверждает, что желтые отметины возникают на голове и в определенных местах туловища, характерных для экспрессии варианта s
ps
p. Однако, если желтизна настолько генерализована, более логично предположить, что осветление происходит за счет гена c
ch(A
y-c
chили c
ch-ee).
Манчестерский терьер
У манчестерского терьера типичный чепрачный окрас. Генотип: a
ta
t
Ньюфаундленд
Для ньюфаундленда допустимы два окраса. Это черный и кофейный (шоколадный). Витни описал голубых и рыжих животных, но они крайне редки. Ландзир — это пегий с крапом ньюфаундленд. Шерсть длинная.
Окрас — Генотип
Черный — A
s-B-D-S-
Кофейный — A
s-bbD-S-
Голубой — A
s-B-ddS-
Ландзир — A
s-B-D-s
p-
Пинчер
Порода имеет самые различные окрасы; красный, черный, черно-подпалый. шоколадный, голубой и голубо-подпалый.
Окрас — Генотип
Черный — A
s-B-D-
Шоколадный — A
s-bbD-
Голубой — A
s-B-dd
Черно-подпалый — a
ta
tB-D-
Шоколадно-подпалый — a
ta
tbbD-
Голубо-подпалый — a
ta
tB-dd
Рыжий (черная мочка носа) — A
y-B-D-
Рыжий (коричневая мочка носа) — A
y-bbD-
Палевый (серая мочка носа) — A
y-B-dd
Палевый (осветленная мочка носа) — A
y-bbdd
Пойнтер
Обычно, пойнтер — это белая собака с окрашенными пятнами, разбросанными по всему телу. Пятна могут быть черными, кофейными или рыжеватыми. Предположительно, что рыжий окрас образуется за счет гена е (Литтл, 1957) Если это так, то генотипы пойнтера можно описать следующим образом:
Окрас — Генотип
Черно-пятнистый — A
s-B-E-s
ps
p
Кофейно-пятнистый — A
s-bbE-s
ps
p
Рыже-пятнистый (черная мочка носа) — A
s-B-ees
ps
p
Рыже-пятнистый (коричневая мочка носа) — A
s-bbees
ps
p
Однако, у некоторых рыже пятнистых собак можно обнаружить следы маски и сероватые уши, что характерно скорее для A
y, чем е. Если это так, то для этих, особей в списки генотипов A
s- следует заменить на A
y- и ее на Е.
Характер пятнистости типичен для s
p. Однако, Литтл (1957) обнаружил, что у пойнтеров США белые ареалы занимают большую площадь, чем у европейских. Поэтому он предположил, что в первом случае собаки имеют не s
ps
p, a s
ws
w.
У некоторых пойнтеров белые ареалы чистые, но у большинства они покрыты крапом соответствующего окрасу цветом. Крапчатость может варьировать в широких пределам. Символ гена Т должен быть добавлен к перечисленным выше генотипам, если собака крапчатая.
Померанский шпиц
Собаки этой породы могут быть самых различных окрасов: черные, кофейные и голубые различных оттенков, рыжие, соболиные, палевые. Каждый цвет может комбинироваться с пегостью (s
p). Шерсть исключительно длинная, густая, шелковистая.
Окрас — Генотип
Черный — A
s-B-D-ll
Кофейный — A
s-bbD-ll
Голубой — A
s-В-ddll
Рыжий (черная мочка носа) — A
y-B-D-ll
Рыжий (коричневая мочка носа) — A
y-bbD-ll
Соболиный (черная мочка носа) — A
y-B-D-ll
Соболиный (коричневая мочка носа) — A
y-bbD-ll
Палевый (серая мочка носа) — A
y-В-ddll
Палевый (осветленная мочка носа) — A
y-bbddll
Пудель
С первого взгляда пудель кажется наиболее представительной породой в отношении разнообразил окрасов. Пуделя бывают черные, коричневые; голубые, светло-рыжие. На основной окрас накладывается действие гена раннего поселения, который с возрастом осветляет окрас в большей или меньшей степени. Литтл считает, что в образовании рыжего и палевого окрасов участвует скорее ген е, чем A
y. Шерсть длинная и жесткая (или вьющаяся), что создает дополнительное богатство оттенков.
Окрас — Генотип
Черный — A
s-B-D-E-gg
Голубой — A
s-B-ddE-gg
Коричневый — A
s-bbD-E-gg
Лилово-серый — A
s-bbddE-gg
Голубо-серый (темно-серебристый) — A
s-B-D-E-G-
Светло-голубой (светло-серебристый) — A
s-B-ddE-G-
Шоколадно-серый — A
s-bbD-E-G-
Шоколадно-серебристый — A
s-bbddE-G-
Светло-рыжий (черная мочка носа) — -B-D-eegg
Светло-рыжий (коричневая мочка носа) — -bbD-eegg
Палевый (серая мочка носа) — -B-ddeegg
Палевый (осветленная мочка носа) — -bbddeegg
Абрикосовый до палевого, кремового — --D-eeG-
Палевый до полностью белого — --ddeeG-
Мопс
Обычно это черные или оленьи собаки с черной маской. Здесь действуют гены A
sи A
yсоответственно. Серебристо-оленьи особи также имеют черную маску, но шерсть на корпусе светло-палевая. Литтл (1957) утверждает, что осветление желтого пигмента происходит благодаря гену c
ch. Однако, ослабление интенсивности может быть также за счет осветляющих руфус-полигенов.
Окрас — Генотип
Черный — A
s-С-
Олений — A
y-С-
Серебристо-олений — A
y-c
chc
ch
Пиренейская горная собака
Эта порода представлена почти полностью или совсем белыми собаками, что является характерным примером проявления крайней степени пятнистости (s
w). Если у собаки есть пятна, то они всегда располагаются только на голове, ушах и крупе. В соответствии с данными Литтля (1957) только 24% собак этой породы полностью свободны от пигментированных участков шерсти.
Окрас пятен зависит от генов основных окрасов, замаскированных белым. Так, волче-серые пятна различных оттенков образуются за счет гена a
sa, действие которого модифицируется осветляющими, полигенами. Лимонные пятна образуются благодаря гену A
y, также при участии руфус-полигенов.
Шерсть густая и длинная, генетически ll. Генотипы большинства цветовых вариаций могут быть представлены как a
sa-lls
ws
wи A
y-lls
ws
w.
У белых пиренейских горных собак с черными глазами ген крайней степени белой пятнистости находится в гомозиготном состоянии (s
ws
w) и полностью подавляет развитие какого-либо пигмента шерсти.
Родезийский риджбек
Очень интересная собака, у которой волосы вдоль спины растут в направлении, противоположном росту остальных волос, т.е. вперед в виде гребня, что и дало название породы. Гребней два. Они должны быть четко обозначены, симметричны и сужены к хвосту. Гребень наследуется рецессивно. Ген, отвечающий за его развитие обозначен, как ds. Окрас желто-красный, обычно с черной маской. Генотип: A
y-dsds
Ротвейлер
Ротвейлер имеет типичный черно-подпалый окрас (a
ta
t). Интенсивность подпалов варьирует, но не так значительно, как у других пород.
Колли
Порода имеет ограниченное число окрасов. Это Ay-рыжие собаки по соболиному типу. Соболиный окрас может быть затенен в различной степени. Встречаются черно-подпалые колли. Мраморные особи генотипически гетерозиготны по гену мерль (Mm). Мраморность трудно выявить у рыжих особей, даже затененных, но она легко определяется у темно-соболиных и черно-подпалых в виде голубо-серых пятен на черном фоне.
Типичная ирландская пятнистость (по типу «воротниковой») колли определяется геном s
i, а не s
p, т.к. белые отметины слишком правильно расположены. Литтл отмечал, что иногда встречаются колли с ослабленным окрасом, т.е. светло-желтые, голубовато-соболиные и голубо-подпалые. Шерсть длинная и ниспадающая; особенно на плечах и груди. У колли описаны припудривающие гены (pp) и CN — ослабление. Но они крайне редки и собаки, несущие их, составляют слишком незначительную часть породы.
Окрас — Генотип
Соболиный — A
y-D-llmms
i-
Черно-подпалый — a
ta
tD-llmms
i-
Соболе-мраморный — A
y-D-llMms
i-
Черно-подпало-мраморный — a
ta
tD-llMms
i-
Голубо-соболиный — A
y-ddllmms
i-
Голубо-подпалый — a
ta
tddllmms
i-
Голубо-соболиный мерль — A
y-ddllMms
i-
Голубо-подпалый мерль — a
ta
tddllMms
i-
Салюки
Собаки самых разнообразных окрасов. Это черный, черно-подпалый, соболиный, рыжий и палевый, сопряженные с пятнистостью и крапчатостью. Литтл (1957) предположил, что в породе встречается ген е. Шерсть гладкая и шелковистая. Встречаются две вариации: с обильной каймой и без нее. Последние, возможно, короткошерстные за счет L, тогда как первые несут гены ll, но с минимальной экспрессией по туловищу. Предполагается, что осветленные палевые собаки несут гены A
yи c
ch, и/или осветляющие руфус-полигены.
Окрас — Генотип
Черный — A
s-S-tt
Черно-подпалый — a
ta
tS-tt
Соболиный — A
y-S-tt
Рыжий — A
y-S-tt
Черно-белый — A
s-s
p-tt
Триколор — a
ta
ts
p-tt
Рыже-белый — A
y-s
p-tt
Черно-белый с крапом — A
s-s
p-T-
Самоед
Отсутствие любого пигмента в шерсти у этих темноглазых собак обусловлено, по-видимому, действием гена s
w. В соответствии с Литтлем (1957) приблизительно 9% самоедов имеют небольшие пигментированные пятна, занимающие не более 10% поверхности туловища. Это говорит о том, что по крайней мере в США описанные Литтлем собаки имели s
ws
w.
Предположено, что самоеды имеют генотип A
s-ees
ws
w. Кроссы самоедов с волче-серыми собаками, гетерозиготными по a
t, давали черно-белых собак. Кроссы с собаками, несущими E
brдавали тигровых и палевых собак. Черные полосы тигровых собак были нормально пигментированы, тогда как желтый фон был очень осветлен, что говорит о наличии осветляющих руфус-полигенов.
Некоторые авторы склоны выделить для окраса самоеда отдельную аллель из альбино-серии. Но это мало вероятно. Наиболее явным кандидатом можно рассматривать узко выявленный аллель шиншилла c
ch. Генотип тогда можно предоставить следующим образом: c
chc
chee или ее с соответствующими руфус-полигенами.
Шерсть длинная, плотная (ll). Генотип: A
s-eells
ws
w.
Шнауцер
В породе допустимы только два окраса. Это сплошной черный и перец с солью. Возникновение последнего загадочно. Предполагается, что это модифицированный чепрачный окрас. Интенсивность желтой пигментации крайне ослаблена, что делает подпалы серыми. Шерсть длинная и жесткая, что затрудняет определение окраса. Генотип черных собак скорее всего A
s-, а собак окраса перец с солью — a
sa.
Скотч-терьер
Возможно, что тигровый окрас скотча сходен с окрасом керн-терьера. При наличии длинной жесткой шерсти действие гена E
brпроявляется для человеческого глаза как смесь черных и желтых волос. Глубина прокрашивания желтых волос варьирует от рыжего до светло-палевого под действием полигенов. Ослабление А
у-рыжего до пшеничного свидетельствует о действии тех же генов на сплошной рыжий. Тигровость варьирует от темной до светлой независимо от интенсивности фона, а только от соотношения количества черных и желтых волос. Наиболее темные тигровые особи выглядят как черные. Тем не менее, нельзя исключить в породе и присутствие гена A
s. Пока это не выяснено.
Окрас — Генотип
Тигровый — A
y-E
br-llWh-
Пшеничный — A
y-E-llWh-
Силихэмтерьер
Белая шубка этого небольшого терьера обязана аллелю экстремальной степени пятнистости s
w. Доказательством этому служит периодическое появление пигментированных пятен в области головы и ушей. Данные Литтля (1967) показывают, что 60% собак пятнисты. Литтл утверждает, что пятна эти настолько малы, что определение скрытых окрасов и кодирующих их генов затруднено. Тем не менее, можно предположить; что генотипы окрасов силихэм-терьера следующие:
A
y-lls
ws
wили a
sa-lls
ws
w.
Шетландская овчарка (шелти)
Генетика окрасов и качества шерсти этой грациозной собаки та же, что у колли (см. выше).
Скай-терьер
Литтл (1957) уделил этой породе большое внимание, однако не пришел к окончательным выводам, кроме того, что генетика окрасов этой породы, возможно, схожа с генетикой керн-терьера (см. выше). Он утверждает, что некоторые индивидуумы имеют осветленную верхнюю часть головы подобно бедлингтонам. Возможно, что осветленные вариации обязаны гену раннего поседения G.
Малый мюнстерлендер
У этой спаниелеобразной собаки на белом фоне разбросаны кофейные пятна. Белые ареалы покрыты крапом, до такой степени, что собака выглядит чалой. Шерсть длинная. Генотип: A
s-bblls
p-Т-.
Гладкошерстная колли
Генетика окрасов идентична генетике колли.
Гладкошерстный фокстерьер
Наиболее обычными для этой породы следует считать черный и чепрачный окрасы. В последнем случае чепрак интенсивно черный и правильной V-образной формы, а подпалы осветлены до золотистого. Присутствует белая пятнистость. Встречаются золотисто-белые собаки. Для них можно предположить: во-первых, экспрессию генов A
yили е; во-вторых, экспрессию гена a
sa, если пигментированные участки расположены только в местах расположения подпалов.
Окрас — Генотип
Черный — A
s-s
p-
Чепрачный — a
sa-s
p-
Мягкошерстный пшеничный терьер
Шерсть длинная, густая и мягкая. Окрас пшеничный, частично из-за действия осветляющих руфус-полигенов, частично благодаря длинной шерсти, которая обнажает более светлый подшерсток. Щенячья шерсть может иметь крап, который с возрастом исчезает.
Генотип: A
y-ll.
Сенбернар
Похоже, что сенбернар имеет соболиный окрас с варьирующим затенением. Неизменно присутствует четко выраженная черная маска. Белая пятнистость «воротникового» типа. Причем расположение отметин является характеристикой породы. Постоянство их расположения говорит о наличии s
is
i. Однако, встречаются более пятнистые собаки, что свидетельствует о наличии в породе гена пегости s
p. Шерсть может быть как короткой, так и длинной. Генотипы этих двух форм: A
y-L-s
i— и A
y-lls
i— соответственно.
Стаффордширский бультерьер
Наиболее распространены черный, рыжий и тигровый окрасы. Время от времени возникают голубые, оленьи и голубо-тигровые особи благодаря наличию в породе гена d.
Окрас — Генотип
Черный — A
s-D-E-
Красный — A
y-D-E-
Тигровый — A
y-D-E
br-
Голубой — A
s-ddE-
Олений — A
y-ddE-
Голубо-тигровый — A
y-ddE
br-
Суссекс-спаниель
Красивые собаки коричневого окраса без белых пятен с длинной шерстью. Генотип: A
s-bbll.
Веймаранер
Эта порода имеет необычный окрас, в стандарте описанный, как серебристо-серый. Такой окрас возникает в результате взаимодействия генов b и d. Известны коротко- и длинношерстные собаки, хотя последние редки. Генотипы:
A
s-bbddL- и A
s-bbddll соответственно. Ген A
s, скорее всего всегда находится в гомозиготном состоянии.
Вельш-корги-кардиган
Эти очаровательные маленькие животные обычно бывают соболиными, тигровыми, черно-подпалыми и голубо-мраморными. Большинство из них пятнисты по типу ирландской (s
i). Литтл (1937) считает, что в породе присутствует также ген s
p, так как встречаются и пегие собаки.
Окрас — Генотип
Соболиный — A
y-E-mm
Тигровый — A
y-E
br-mm
Черно-подпалый — a
ta
tE-mm
Мраморный — a
ta
tE-Mm
Вельш-корги-пемброк
В этой породе встречаются только два окраса: соболиный различных оттенков и черно-подпалый, сопряженные с ирландской пятнистостью.
Окрас — Генотип
Соболиный — A
y-s
i-
Черно-подпалый — a
ta
ts
i-
Вельш-спрингер-спаниель
Основной окрас рыже-белый, шерсть длинная, мочка носа печеночного цвета, что говорит о наличии гена b в гомозиготном состоянии. Литтл считает, что рыжий окрас обязан гену е. Тогда генотип будет bbeells
p-.
Вельштерьер
По окрасу и типу шерсти вельштерьера можно считать маленькой копией эрдельтерьера. Он отличается только более светлым чепраком. Когда осветление чепрака происходит в значительной степени, такой окрас называют гризли. Генотип: a
sa-Wh-.
Вест-хайланд-вайт-терьер
Литтл (1957) считает, что белый окрас этих собак обязан взаимодействию генов А
уи c
ch. При селекции как можно более белых особей генотип A
y-c
chc
chможет давать как бледно палевых, так и чисто белых собак, хотя, казалось бы, что генотип окраса этого терьера должен быть сходен с генотипом окраса силихэм-терьера (см. выше). Однако, Литтл обнаружил у некоторых собак светло-кремовые кончики на белых волосах, тогда как следы пигментации на голове, характерные для s
ws
wвстречаются крайне редко. Однако, отсутствие таких индивидуумов в породе возможно, если селекция, направленная на получение белого окраса при генотипе s
ws
wпрошла наиболее успешно. Шерсть грубая и длинная (ll).
Уиппет
Уиппеты — это миниатюрные грейхаунды и генетика у них схожа (см. выше).
Жесткошерстный фокстерьер
Окрасы идентичны окрасам гладкошерстного фокстерьера (см. выше). Шерсть жесткая.
Окрас — Генотип
Черный — A
s-s
p-Wh-
Чепрачный — a
sa-s
pWh-
Йоркширский терьер
Окрас чепрачный необычных оттенков. Чепрак покрывает большую часть туловища. У щенков он более темный и светлеет с возрастом. Литтл (1957) предположил что такое явление обязано гену G. Возможно также, что возрастное осветление обязано длинным прямым волосам, которые обнажают более светлый голубой подшерсток.
Рисунок 1
Экспрессия аллелей белой пятнистости показана в десяти степенях. Степень 0 является показателем того, что собака имеет незначительную пятнистость или вовсе ее лишена. Степень 10 — собака полностью белая (на рисунке не отражено).