 |
|
Популярные авторы:: Борхес Хорхе Луис :: БСЭ :: Азимов Айзек :: Станюкович Константин Михайлович :: Грин Александр :: Лесков Николай Семёнович :: Раззаков Федор :: Чехов Антон Павлович :: Андреев Леонид Николаевич :: Нортон Андрэ Популярные книги:: Дюна (Книги 1-3) :: Синее платье :: Крылатый человек :: Быть драконом :: The Boarding House :: Ты, Марианна :: Ее пятый муж? :: Возвращение "Чайки" :: Утро космоса. Королев и Гагарин :: Мармелад.RU |
Аксиомы биологииModernLib.Net / Биология / Медников Борис Михайлович / Аксиомы биологии - Чтение (стр. 10)
Иногда же ответов несколько, причем они отнюдь не равноценны. Вот хороший пример. Представьте разреженный лес или саванну, где трава выжигается солнцем. У крупных копытных в этой зоне есть хороший источник питания – листва деревьев. Но как до нее добраться? Самый прямой ответ – отбор на увеличение размеров тела. По этому пути пошли гигантские безрогие носороги олигоцена – индрикотерии. Нельзя сказать, чтобы решение было удачным. Ведь если рост увеличивается, то вес также возрастает пропорционально третьей степени роста. Размеры возросли вдвое, а расходы на питание – в восемь раз. 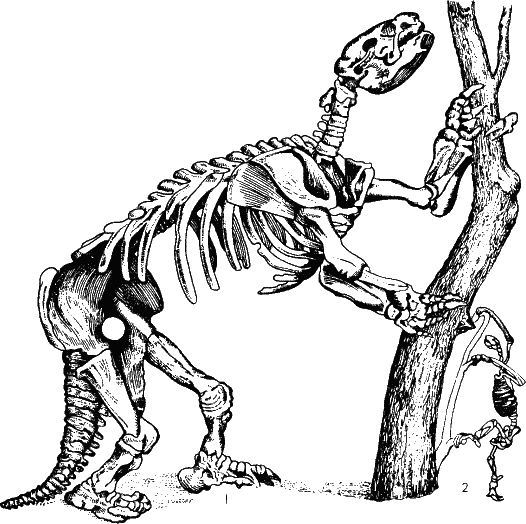
Рис. 45. А вот еще один способ добраться до листьев на деревьях. Гигантские южно-американские ленинцы поднимались на дыбы и подгибали дерево, охватывая ствол передними лапами с мощными когтями. На рисунке скелет ленинца – милодона, раскопанный в Аргентине еще Чарлзом Дарвином, справа (2) скелет современного ленинца. По-видимому, так же вели себя халикотерии, о которых упоминалось выше в связи с ошибкой Кювье. Другие животные вставали на задние ноги, а передними (нередко с мощными загнутыми когтями) подгибали ветки с листьями и плодами. Так вели себя, очевидно, халикотерии, о которых уже упоминалось. Предел этой тенденции – гигантские ленивцы, мегатерии, вымершие в Южной Америке сравнительно недавно, может быть, не без содействия человека. 
Рис. 43. Как мы уже видели, пропорциональное увеличение размеров тела для того, чтобы достать листья с высоких деревьев, энергетически невыгодно. Большего успеха в жизненной борьбе достигли животные со специализированными приспособлениями. Слон дотягивается до веток хоботом (это продукт разрастания носа и верхней губы), у жирафы удлиняются шея, ноги и язык. Геренук (она же жирафовая газель, или антилопа Уэллера) также добирается до листьев на высоких кустарниках с помощью длинной шеи и ног; к тому же она отлично стоит на задних ногах, балансируя передними. У слона появляется длинный хобот, с помощью которого он дотягивается до листвы. Но, по-видимому, наиболее перспективный ответ – удлинение шеи. У всем известной жирафы длинная шея и длинные ноги, что позволяет ей объедать листву на высоте, для других копытных недоступной. К этому еще добавляется длиннейший язык (жирафа и ее родственница окапи могут облизывать себе уши). 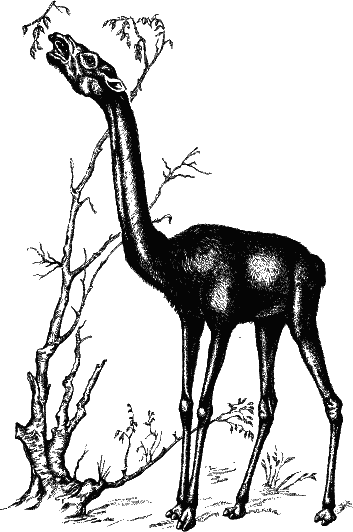
Рис. 44. Внешний вид жирафы известен каждому. Но не каждый знает, что в миоценовом периоде в Северной Америке обитали жирафоподобные верблюды – альтикамелюсы (на рисунке реконструкция внешнего вида). По-видимому, жирафоподобный тип строения оказался очень удачной эволюционной находкой, если повторялся по меньшей мере два раза. И, однако, альтикамеллос остался верблюдом, на жирафу он похож только внешне. Этот удачный тип строения был использован эволюцией не один раз. В миоценовом периоде в Северной Америке обитали жирафоподобные верблюды – альтикамелюсы. А в Сомали, в самых жарких и бесплодных местностях, и сейчас благоденствует удивительная жирафовая газель (она же геренук, или антилопа Уэллера). У нее длинная шея и ноги; кроме того, геренук может объедать листья на высоком кустарнике, стоя на задних ногах. 
Рис. 48. Животные, обитающие на деревьях, также дают нам яркие примеры конвергенции. Сходный образ жизни приводит к возникновению сходных приспособлений. Слева – хамелеон, справа – цейлонская древолазящая ящерица агама. Их связывает весьма отдаленное родство, но внешний вид сходен. Этого мало: многие детали структуры, например охватывающая ветвь конечность, возникают и у пресмыкающихся и у млекопитающих (внизу скелет конечности хамелеона и лапа кус-куса – древолазящего сумчатого, обитающего на Новой Гвинее). Есть и лягушки с подобными лапами. Вот такие примеры конвергентного, независимого возникновения сходных признаков до сих пор многим кажутся убедительным доказательством направленности эволюционного процесса. Причем направленным считается даже не действие отбора, с чем можно согласиться, а сама изменчивость. Самые последовательные из сторонников такого толкования полагают, что абсолютно идентичные формы могут возникать из несходных, что один вид может иметь двух, а то и более предков. Это равносильно утверждению, что первоначально несходные последовательности ДНК в результате накопления мутаций станут идентичными и приведут к возникновению идентичных фенотипов. В свете наших сегодняшних знаний о структуре ДНК и механизме ее генетических изменений подобная декларация не заслуживает опровержения. Вероятность того, что два романиста из разных стран независимо друг от друга напишут одинаковые до последней запятой романы, куда выше вероятности конвергентного возникновения видов. 
Рис. 47. Один из популярнейших примеров конвергенции – глаз осьминога (А) и глаз человека (Б). Одинаковыми цифрами показаны аналогичные части. Казалось бы, сходство идеальное (у осьминога хрусталик более округлый из-за того, что у воды больший коэффициент преломления, чем у воздуха). Но глаз осьминога «наводится на фокус» приближением или удалением хрусталика от сетчатки, как объектив фотоаппарата. У человека фокусное расстояние изменяется путем изменения кривизны самого хрусталика. Несмотря на внешнее сходство, принципы работы этих оптических приборов совершенно различны. Еще одно следствие, вытекающее из четырех наших аксиом: конвергентными могут быть только признаки, части структур, но не целые структуры, а тем более не сами организмы. Много пишут об удивительном конвергентном сходстве глаза человека и осьминога. Внешне они действительно похожи, вплоть до выражения. Но принципы действия совершенно различны. Глаз осьминога, как объектив фотоаппарата, «наводится на фокус» приближением или удалением к сетчатке. У человека фокусное расстояние регулируется изменением кривизны самого хрусталика (представьте оптический прибор с эластичной линзой, фокусное расстояние которой изменяется, например, при накачке в линзу жидкости, сильно преломляющей свет; насколько я знаю, подобный принцип в технике не был использован). Сумчатый крот, обитающий в Австралии, удивительно похож на нашего крота, но у него имеется сумка для вынашивания детенышей и задние концы нижней челюсти, как у всех сумчатых, загнуты внутрь. Это вполне понятно: отбор приводит к конвергентному сходству по признакам, которые полезны в конкретных условиях (например, к подземному образу жизни), но оставляет те, которые могут быть разными без нарушения их функций. Поэтому на Земле нет двух видов с одинаковыми генетическими программами. 
Рис. 49. Конвергенция может сыграть злую шутку с исследователем. Что изображено на этом рисунке? Кроты, скажете вы и будете правы. Только это не наши обычные кроты: вверху изображен златокрот из Южной Африки, тоже насекомоядное, но из другого семейства, а внизу сумчатый австралийский крот. Еще раз подчеркнем: конвергентные формы – это книги разного содержания в одинаковых переплетах. Этого мало: можно утверждать, что каждый вид уникален и, раз исчезнув с лица Земли, не возникает вновь. Нельзя воссоздать заново мамонта и Стеллерову корову, среднеазиатского тигра и гепарда, вымерших в последние годы. Даже когда мы научимся сами создавать новые виды, мы можем сделать их лучше вымерших, но идентичных не получим. Потеря каждого вида безвозвратна. Возможности человека уничтожить любой вид, включая себя, безграничны, возможности исправить содеянное равны нулю. И дело тут не в ограниченности наших знаний, а опять же во втором начале термодинамики, запрещающем воссоздать утерянную генетическую программу. 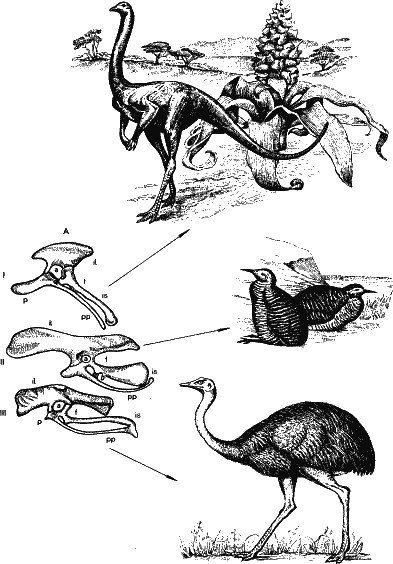
Рис. 50. Еще раз напомним: конвергентными могут быть лишь детали строения организма, но не сами организмы. Здесь изображены тазовые кости динозавра (I), американской птицы тинаму (II) и американского страуса нанду (III). Сходство действительно большое. Но даже самого птицеподобного динозавра струтиомимуса (само название его – подражающий страусу) нельзя спутать ни с тинаму, ни со страусом. Так же различаются тинаму и нанду. Один и тот же болт может быть использован и в самолете и в автомобиле, но эти структуры легко различаются. Обратное положение из этого тезиса: если в геномах двух видов найдется хотя бы одна нуклеотидная последовательность достаточной (более 50 нуклеотидных пар) протяженности, они связаны родством, ведут начало от одного предка. Ибо вероятность возникновения одной последовательности из двух первоначально несходных слишком мала, чтобы ее можно было принимать во внимание. С большей вероятностью вода, выплеснутая из стакана, сама, самопроизвольно соберется в него. На этом принципе основана новая отрасль систематики – геносистематика, основы которой заложил академик А. Н. Белозерский. И не только геносистематика. Рискну высказать утверждение, что геносистематика, теория эволюции геномов и т. д. в конце концов окажутся лишь частью общей теории эволюции последовательно реплицирующихся систем. Такой теории еще нет. Однако и сейчас можно полагать, что все системы, передающиеся из поколения в поколение с накоплением ошибок репликации, под контролем внешней среды изменяются, подчиняясь сходным закономерностям. Под эту категорию попадают не только объекты биологической эволюции, но и, например, человеческие языки, обычаи, обряды, мифы (включая религии), сказки и многое другое. Когда-нибудь я напишу и об этом. Заключение Нет ничего труднее, чем завершение книги. С каждой страницей, приближающей к концу, умножаются сомнения: обо всем ли написал и так ли, как нужно. Воистину, по словам Гёте: «Alles war gesagt, doch alles bleibt zu sagen!» — «Все было сказано, лишь все сказать остается!» К счастью, на сей раз положение лучше. Вариант книги в 1980 году был опубликован популярным журналом «Наука и жизнь»; как водится, в редакцию пошли письма имеющих досуг читателей, и теперь ясно, что я недоучел в работе. Разумеется, далеко не все письма могли быть использованы. Редакция предпочла сама отвечать на те письма, которые не содержа ли ничего, кроме брани. Однако и среди тех, что были мне направлены для ознакомления, находились такие, которые до сих пор меня ставят в тупик. Биология наряду с науками, являющимися ее практическим приложением (медициной, агробиологией и т. д.), в представлении многих моих корреспондентов кажется принципиально иной отраслью знания, чем, скажем, машиностроение или теория атомного ядра. Вряд ли кому-нибудь придет в голову усомниться в фактах и выводах, изложенных, к примеру, в статье об управляемом термоядерном синтезе. А вот биология, особенно общая – здесь «каждый мнит себя стратегом, видя бой со стороны». И вот бедняга-автор, рискнувший ступить на трудную стезю популяризации, получает письма об установлении связи с «космической цивилизацией Эридана Тулона Элсилон», о «всеобъемлющем законе энергии разности потенциалов» и т. д. и т. д. В эпитетах корреспонденты не стесняются, слова «идеализм», «недомыслие», «безграмотность», «узкая щель мирозрения» (так! – Б. М.) на общем фоне звучат чуть ли комплиментами. Не скажу, чтобы подобные письма было приятно читать. Однако и из них можно почерпнуть полезное, чтобы еще раз остановиться на тех вопросах, о которых мои корреспонденты имеют ложное представление. Любопытно, что читателей, насколько могу судить, в первую очередь интересуют именно философские проблемы, которые возникают при попытках аксиоматизировать биологию. Много говорится, например, о законе причинности. По-видимому, все довольно легко согласились с тем, что учение Ламарка этот закон нарушает и мы должны сделать выбор: или мираж направленной приспособительной изменчивости, или же сохранение одного из основных принципов современного естествознания. Однако для некоторых мираж оказывается предпочтительнее закона причинности. В некоторых письмах категорично утверждается: закон причинности устарел как для живой природы, так и для неживой. Один мой корреспондент упоминает тахионы, которые будто бы движутся со сверхсветовой скоростью, и настоятельно рекомендует мне прочесть популярные брошюры на эту тему. (Невольно вспоминается фарадеевское: «Популярные книги ничему научить не могут». Начинать можно с них, но завершать обучение, если дело касается физики, например, лучше все-таки вузовским курсом Ландау и Лифшица.) Тахионы – частицы гипотетические, экспериментально они не обнаружены, реальное существование их не доказано, и поэтому опровергать с их помощью принцип причинности, мягко выражаясь, преждевременно.[10] Не меньшее раздражение вызвало у ряда читателей то, что я подчеркиваю везде функциональную подоплеку аксиом, не связывая жизнь с каким-либо химическим веществом. В этом усматривали идеализм и «богословную мистику». Однако вынужден еще раз подчеркнуть: да, в конкретных условиях нашей Земли материальным субстратом генетических программ оказались нуклеиновые кислоты, а функциональным, эффекторным субстратом – белки. Но из этого отнюдь не следует, что везде во Вселенной дело обстоит точно так же. Даже в условиях, приближающихся к земным, в том интервале давлений и температур, в котором существует жидкая вода, возможны материальные субстраты жизни, которые нам сейчас показались бы невероятными. Тем более это относится к планетам, покрытым морями, например, из жидкого метана и аммиака, даже из жидкой серы. Как бы нам ни казалось невозможным появление и эволюция жизни в таких условиях, вероятность этого во Вселенной нельзя отрицать с порога. Сможем ли мы узнать жизнь, столь непохожую на земную, или же будем искать в космосе только собственное отражение? Не придется ли нам тогда искать слишком долго? Когда космонавты будущего обнаружат где-то во Вселенной структуру, пусть любого, самого фантастического вида и строения, если эта структура может поддерживать свою целостность и воспроизводить себе подобных согласно вложенной в нее программе, реплицирующейся матричным способом, они должны признать ее живой. Дело даже не в будущих космонавтах, – понять, что такое жизнь, мы должны уже сейчас. Ибо это означает познание самого себя. Пожалуй, наибольшее разнообразие можно было обнаружить в определениях понятия жизнь». Каких только формулировок не предлагают в дискуссиях и в письмах! Как правило, они не выдерживают критики, мягко скажем, из-за недостаточной информированности авторов. Приведу лишь один пример, из самых характерных. Один автор чрезвычайно агрессивно отстаивающий свою точку зрения, доказывает, что в понятие жизни должен входить активный поиск и захват источников энергии. По его мнению, жизнь начинается с амебы: ведь она активно захватывает пищевые частицы. А вот бактерии, грибы, да и растения – неживые. Ясно, автор не подозревал о существовании хотя бы бактерии бделловибрион бактериоворус (что: означает пиявкообразный вибрион, пожирающий бактерий). Этот маленький вибрион нападает на других бактерий, причем движется с огромной скоростью – до ста своих длин в секунду! Если бы человек в своих масштабах мог двигаться так быстро, он пробегал бы стометровку за долю секунды. Бделловибрион ударяет бактерию-жертву с такой силой, что она по инерции проходит немалое расстояние. А крошечный хищник тем временем стремительно просверливает клеточную стенку жертвы (скорость вращения свыше ста оборотов в секунду), за несколько секунд проникает в пространство между стенкой и мембраной, и через четыре часа из «съеденной» изнутри бактерии выходят новые, молодые вибрионы. Если уж это не активный поиск источника энергии… Кстати, бделловибрионы – не единичный каприз природы. Хищные бактерии – мощный фактор очищения воды. Не слыхивал мой оппонент и о хищных грибах, обитающих в почве, мицелий которых, переползая с места на место, захватывает мелких животных; говорят уже об их применении для борьбы с почвенными нематодами – вредителями сельского хозяйства. Но неужели мой оппонент ничего не знал о насекомоядных растениях, хотя бы о всем известной росянке, которую изучил еще Чарльз Дарвин? Или о часто встречающейся в наших водоемах пузырчатке: достаточно мелкому животному прикоснуться к чувствительному «волоску», запоры ловчего пузырька резко открываются, и ток воды затягивает жертву внутрь, где она и переваривается. Примерно так же захватывает добычу рыба – морской черт. От растений, питающихся как животные, перейдем к животным, питающимся как растения. Симбиотические одноклеточные водоросли – зоохлореллы и зооксантеллы – синтезируют углеводы в клетках корненожек и гидр, губок и кораллов, плоских червей и моллюсков. О том, что такой способ питания (без активного поиска и захвата пищи) весьма эффективен, свидетельствуют хотя бы протянувшиеся на тысячи километров коралловые рифы. Ограничимся двумя примерами. Осыхающая во время отлива поверхность песчаных пляжей Северного моря покрывается оливково-зелеными пятнами. Это выползают на поверхность песка, под солнечные лучи маленькие плоские черви – конволюты. Тело их заполнено симбиотическими одноклеточными водорослями – зоохлореллами. За время отлива симбионты успевают насинтезировать углеводов и для себя, и для переваривающего их черня. Растение это или животное? Гигантский двустворчатый моллюск коралловых рифов – тридакна, если никем не потревожена, открывает свои створки и выворачивает навстречу солнечным лучам оливково-бурую мантию. Поверхность мантии как бы шагреневая. Она покрыта тысячами мелких выступов, заканчивающихся линзоподобными структурами из прозрачных, хрящеподобных (гиалиновых) клеток. Каждый такой бугорок – настоящий световод, собирающий солнечные лучи и передающий их внутрь ткани, к скоплениям симбиотических водорослей – зооксантелл. У тридакны недоразвита пищеварительная система, она живет главным образом за счет своих симбионтов, потребляемых клетками-фагоцитами. Везде, где активное питание можно заменить пассивным, за счет утилизации энергии солнечных лучей, животные охотно делают это. И наоборот, там, где в почве мало азота, растения приобретают способность к хищничеству. Мы видим, что построения нашего автора, строгого хулителя дарвиновской теории эволюции и современной генетики, рассыпаются как карточный домик при малейшем соприкосновении с фактами. Ибо это порождения – не постесняемся резкого слова – элементарного невежества. Вряд ли имело бы смысл останавливаться на явлениях подобного рода, если бы это невежество не было воинствующим. Слов нет, недостаток информации никогда не приносил пользы, однако воинствующее невежество может принести огромный вред, чему мы видим немало примеров в прошлом. Ведь тот же автор и его единомышленники голословно отрицают роль ДНК в явлениях наследственности. И это сейчас, когда успехи генной инженерии убедительно показывают, что до широкого внедрения в практику методов направленного изменения наследственности остаются считанные годы. Генная инженерия может произвести подлинную революцию в медицине и сельском хозяйстве, промышленности и охране природы. (В то же время не будем закрывать глаза: в руках недобросовестных людей она может быть не менее страшной, чем ядерное оружие.) Зачеркивать достижения биологии последних лет – все равно что отрицать существование атомов в начале августа 1945 года[11]. И самое малое, что мы можем сделать, для того чтобы не отставать в развитии биологии, – решительно пресекать все попытки подобного, как писали Ильф и Петров, «головотяпства со взломом». Но довольно об этом. Еще раз перечислю выдвинутые в этой книге в качестве аксиоматических положения, лежащие в основе жизни. Именно: 1) живые организмы состоят из фенотипа и генотипа, то есть генетической программы; 2) генетические программы не возникают заново, а реплицируются матричным способом; З) в процессе репликации неизбежны ошибки на микроуровне, случайные и непредсказуемые изменения генетических программ; 4) в процессе постройки фенотипа эти изменения многократно усиливаются, что делает возможным селекцию единичных квантовых событий на макроуровне. Осмелюсь утверждать, что этих положений достаточно (как говорят математики: необходимо и достаточно) для объяснения всех феноменов живой природы – от возникновения первого сократимого волокна или первой молекулы фермента до становления мыслящего мозга. Я предвижу хор возмущенных голосов, основным лейтмотивом которого будет: «Нельзя сводить биологию к физике и химии!» Но разве рассмотренные в этих очерках аксиомы характерны для неживой природы? Где же тут сведение? Появилось уже хлесткое определение «редукционизм» так называют направление, которое не прибегает для объяснения сущности жизни к аналогам «жизненной силы». Но, как я уже писал вначале, мы не сводим биологию к химии и физике. Мы выводим биологию из химии и физики. Это не редукционизм, а дедукционизм (различие в одной букве и во всем смысле термина). Если уж говорить о редукционизме, то только в том смысле, что все эти положения прямо вытекают из положений физики и химии и только в этом понимании «сводятся» к ним. Ибо что значит – объяснить? Это и есть свести сложное явление к более простым, для нас уже понятным, объясненным. Утверждение, что это-де отрицает «специфику жизненных явлений», по сути дела, повторяет анекдотическое определение жизни, цитированное мной раньше («живое состоит из живых объектов, неживое – из неживых»). Естествоиспытатель в наши дни обязан быть редукционистом, иначе он рискует остаться просто болтуном. Вот, например, что пишет о редукционизме советский философ Р. С. Карпинская: «…если вопрос о редукционизме рассматривать в методологическом плане, то сущностью «сведения» сложных биологических процессов к более простым является обнаружение на молекулярном (или ином, более простом по отношению к изучаемому) уровне таких фундаментальных характеристик, которые при их теоретическом обобщении позволят сформулировать некие абстрактные понятия, выступающие начальным пунктом движения познания «вверх», ко все более сложным уровням биологической организации. Эти понятия должны работать на всех уровнях, наполняясь все более конкретным, все более богатым содержанием» (курсив мой. – Б. М.). Прошу извинения у читателей за длинную цитату: на мой взгляд, в ней удачно подчеркивается необходимость неких абстрагированных заключений (которые я назвал аксиомами, но не в слове суть: назовите их постулатами, принципами, положениями, в конце концов, русский язык достаточно богат). Кроме того, формулируются условия, которым должны соответствовать наши аксиомы. 1. Хотя они (аксиомы) выводятся на молекулярном уровне (деление организма на генотип и фенотип, матрицирование генотипа, ошибки матрицирования, усиление ошибок, делающее их доступным отбору), с их помощью можно познать более сложные уровни биологической организации. 2. Они должны быть всеобщими, проявляться на всех уровнях и во всех случаях. Частные закономерности должны выводиться из них, как теоремы из аксиом. И включение в систему аксиом «лишней», не работающей на всех уровнях, такая же, если не худшая, ошибка, как включение неверной, не обоснованной фактами и противоречащей принципам физики и химии. С этой точки зрения рассмотрим вопрос: достаточна ли наша система аксиом, не следует ли чего-либо добавить к ней? Признаюсь, что для меня этот вопрос проклятый, я ломал голову над ним годами. Остановлюсь для примера на положении, которое чуть было не стало пятой аксиомой. Его можно было бы выразить двумя словами: генетическая рекомбинация. Генетическая рекомбинация – обмен частями генетических программ – возникла, по-видимому, на чрезвычайно ранней стадии становления жизни. У самых простейших организмов – бактериофагов – описан сложный процесс обмена кусками ДНК. После заражения бактериальной клетки фагом в ней образуется 30—50 копий молекул ДНК, реплицированных с материнской молекулы, впрыснутой в бактерию. Дочерние молекулы могут спариваться, сближаться гомологичными частями; затем в зонах спаривания происходят разрывы нитей ДНК, обмен скрещенными участками и последующая сшивка. Все это очень напоминает хорошо известный генетикам перекрест хромосом у высших организмов – кроссинговер, при котором хромосомы обмениваются гомологичными частями. В результате, если в одной молекуле ДНК произошла одна мутация, а в другой – другая, они могут объединиться в генетической программе фага и совместно попасть под действие отбора. Скорее всего, механизм этот развился из репарации, о которой мы здесь говорили. Репарационные ферменты – лигазы – сшивают разорванные жестким излучением или иным мутагеном нити ДНК, восстанавливают целостность генетической программы. А так как в одной клетке несколько десятков фаговых ДНК, то высока вероятность объединения частей разных молекул. Рекомбинация оказалась выгодной, допускающей объединение генетических изменений у разных организмов. У бактерий перенос генетической информации осуществляется от клетки к клетке непосредственно кусками ДНК (трансформация), в результате изучения этого процесса и была доказана роль ДНК как субстрата наследственности. Часто гены переносятся от бактерии к бактерии фагами, фаг может прихватить кусок ДНК хозяина и передать его в новом поколении другому. Этот процесс называется трансдукцией. Обычно ее осуществляют плазмиды, «ручные» фаги, ставшие симбионтами бактерий и не убивающие хозяина интенсивным размножением. Из подобных механизмов у бактерий в конце концов развился оригинальный половой процесс конъюгация. При нем клетки соприкасаются, между ними образует цитоплазматический мостик по которому нить ДНК перетекает из одной бактерии в другую. Любопытно, что для передачи генного материала бактерия должна иметь внехромосомный генетический элемент, названный фактором F, который, возможно, потомок плазмиды. Не имеющие его штаммы (F – ) могут только принимать чужую ДНК (аналогия между самками и самцами у высших животных; кстати, «самцов» то есть имеющих фактор F, довольно мало: у кишечной палочки их в десять раз меньше, чем «самок»). Плазмида, ведущая происхождение от фага, может вообще стать частью бактериальной хромосомы, тогда она называется эписомой. Также и F – фактор, включаясь в геном бактерий, повышает их способность к конъюгации в тысячу раз. Из школьного курса общей биологии вы должны знать, что генетическая рекомбинация у высших организмов гораздо сложнее. И у них есть кроссинговер. Но в отличие от бактерий геном у них настолько велик, что не может быть объединен в одной хромосоме.[12] Ядерные организмы – эукариоты – имеют в клетках от двух (у малярийного плазмодия и лошадиной аскариды) до тысячи и более хромосом, «томов» генетических программ. Перед каждым делением клетки генетические программы реплицируются. Однако так бывает не всегда. Перед наступлением полового процесса происходит мейоз – редукционное деление. Хромосомы при нем не делятся, а расходятся по клеткам, из которых потом формируются половые. Так, у человека в норме 46 хромосом из них две половые, остальные 44 идентичны у обоих полов (аутосомы). 23 хромосомы человек получает от отца, 23 – от матери, а каково сочетание отцовских и материнских хромосом в яйцеклетке или спермии – это дело случая. Число сочетаний здесь равно двум в степени n – числу хромосом в гамете. Отсюда следует, что, например, у дрозофилы с ее четырьмя хромосомами в гамете число вариантов гамет 24 = 16, у кролика 222 = 4 385 000. Предоставляю читателям самим прикинуть число вариантов гамет у папоротника (количество хромосом в гамете 630). Слияние половых клеток восстанавливает прежнее количество хромосом возникает новая генетическая программах с которой организм входит в жизнь. При смене поколений эти программы рассыпаются чтобы заново возникли другие. Генетическая рекомбинация непрерывно тасует их, как карты, поставляя отбору поистине неисчерпаемый материал. Здесь есть и отрицательная сторона: какая-нибудь сверхудачная комбинация хромосом бесследно исчезает в новом поколении, у гениальных родителей появляются заурядные дети. По-видимому, этот великий по простоте механизм рекомбинации возник в процессе эволюции только один раз. У человека и сосны, инфузории и дрожжевого грибка половые ядра, сливающиеся впоследствии, возникают одинаковым путем: ядро с нормальным набором хромосом не разделяясь реплицирует их, так что образуется четыре набора генетических программ. Затем клетка (или только ядро у инфузорий) дважды делится, получаются четыре клетки с половинным, гаплоидным набором. Часто, особенно при формировании яйцеклеток, три из них рассасываются, дальнейшее развитие суждено только одной. Лишь немногие панцирные жгутиковые имеют одноступенчатое редукционное деление; у них это, скорее всего, вторичное упрощение. Так, может быть, объявим генетическую рекомбинацию непреложным законом живого, пятой аксиомой? Увы, дело обстоит не столь просто. Многие организмы потеряли способность к генетической рекомбинации и благоденствуют, мы не имеем права исключать их из мира живого. Таковы всем известные амебы и инфузории потерявшие способность образовывать половое ядро, все формы, размножающиеся партеногенетическим путем, без оплодотворения (а их многие тысячи – от простейших до некоторых пород индеек). Наконец, так называемые апомиктические растения, образующие семена из нормальных, диплоидных клеток с двойным набором хромосом, хотя бы одуванчики, каждую весну золотым потопом заливающие наши газоны. Или же растения в принципе способные к рекомбинации, но размножаемые черенками клубнями, отводками (картофель бананы и т. д. и т. д.). Кроссинговер, казалось бы, шире распространен, чем половой процесс, но, например, самцы дрозофил к нему не способны. Отсюда делаем вывод: наличие генетической рекомбинации – не безусловный критерий живого. Это не условие жизни, а ее завоевание, не аксиома, а теорема (правда, одна из самых основных). Мой коллега Алексей Владимирович Яблоков, ознакомившись с рукописью, предложил в качестве пятой аксиомы так называемое «давление жизни», геометрическую прогрессию размножения. Рассмотрим этот вопрос. 1, 2, 3, 4, 5, 6, 7, 8, 9, 10, 11 |
|||||||