При одноячеистой конвекции положение полюсов подъема и опускания вещества всегда будет несколько отличаться от идеального (точно по диаметру планеты); там, где соединяющие их «меридианы» будут самыми длинными, образуется застойная область, в которой вещество не теряет железа и потому постепенно оказывается тяжелее окружающей его среды. Через некоторое время оно «проваливается» вглубь мантии, создавая второй полюс опускания, и превращая конвекцию в двухъячеистую. Двухъячеистая конвекция постепенно ослабляется и затем переходит в одноячеистую (одна из ячеек как бы «съедает» вторую), и конвекционный цикл начинается заново. Таким образом, взаиморасположение континентов определяется фазой конвекционного цикла в мантии – и наоборот: фаза конвекционного цикла, имевшая место в некую геологическую эпоху, может быть определена исходя из взаиморасположения континентов, реконструированного палеомагнитными, палеоклиматологическими и др. методами. Понятно, что все эти изменения весьма существенно влияют на климат соответствующей эпохи, а через него – на функционирование ее биосферы.
Завершив раздел, посвященный эволюции самой Земли, мы приступаем теперь к изучению эволюции жизни на ней. Сразу оговорюсь: я не собираюсь здесь ни углубляться в дебри определений того, что такое «жизнь», ни обсуждать чисто химические аспекты этого явления – это увело бы нас слишком далеко от темы спецкурса
. Наш подход к проблеме жизни на Земле будет сугубо функциональным, и в его рамках нам следует принять одно аксиоматическое утверждение: эволюция биосферы и составляющих ее экосистем идет в целом в сторону возникновения все более совершенных, т.е. устойчивых и экономных, круговоротов вещества и энергии. Совершенствование циклов направлено на то, чтобы минимизировать безвозвратные потери биологических систем: экосистема стремится препятствовать вымыванию микроэлементов и захоронению неокисленного углерода, переводить воду из поверхностного стока в подземный, и т.д. Поэтому с общепланетарной точки зрения жизнь следует рассматривать как способ стабилизации существующих на планете геохимических циклов.
Что же касается происхождения жизни на Земле, то обычно проблему эту, еще со времен Э. Геккеля (1866), сводят к чисто химической задаче: как синтезировать сложные органические макромолекулы (прежде всего – белки и нуклеиновые кислоты) из простых (метана, аммиака, сероводорода и пр.), которые составляли первичную атмосферу Земли. Следует честно признать, что даже эта, в общем-то техническая, задача чрезвычайно далека от своего разрешения. В двадцатые годы А.И. Опарин и Дж. Холдейн экспериментально показали, что в растворах высокомолекулярных органических соединений могут возникать зоны повышенной их концентрации – коацерватные капли – которые в некотором смысле ведут себя подобно живым объектам: самопроизвольно растут, делятся и обмениваются веществом с окружающей их жидкостью через уплотненную поверхность раздела. Затем, в 1953 году, С. Миллер воспроизвел в колбе газовый состав первичной атмосферы Земли (исходя из состава современных вулканических газов), и при помощи электрических разрядов, имитирующих грозы, синтезировал в ней ряд органических соединений – в том числе аминокислоты. Через некоторое время С. Фоксу удалось соединить последние в короткие нерегулярные цепи – безматричный синтез полипептидов; подобные полипептидные цепи были потом реально найдены, среди прочей простой органики, в метеоритном веществе. Этим, собственно говоря, и исчерпываются реальные успехи, достигнутые в рамках концепции абиогенеза – если не считать того, что было ясно осознано по крайней мере одно фундаментальное ограничение на возможность синтеза «живых» (т.е. биологически активных) макромолекул из более простых органических «кирпичиков».
Дело в том, что многие органические соединения представляют собой смесь двух так называемых оптических изомеров – веществ, имеющих совершенно одинаковые химические свойства, но различающихся так называемой оптической активностью. Они по-разному отклоняют луч поляризованного света, проходящий через их кристаллы или растворы, и в соответствии с направлением этого отклонения называются право– или левовращающими; свойством этим обладают лишь чистые изомеры, смеси же их оптически неактивны. Явление это связывают с наличием в молекуле таких веществ так называемого асимметричного атома углерода, к четырем валентностям которого могут в разном порядке присоединяться четыре соответствующих радикала (рисунок 13). Так вот, эти химически идентичные вещества, как выяснил еще в 1848 г. Л. Пастер, вовсе не являются таковыми для живых существ: плесневый гриб пенициллиум, развиваясь в среде из виноградной кислоты, «поедает» лишь ее правовращающий изомер, а в среде из молочной кислоты – левовращающий (на этом, кстати, основан один из методов разделения оптических изомеров), человек легко определяет на вкус изомеры молочной кислоты.
РИСУНОК 13. (пропущен) «Левая» и «правая» молекулы аланина.
Сейчас известно, что все белки на нашей планете построены только из левовращающих аминокислот, а нуклеиновые кислоты – из правовращающих сахаров; это свойство, называемое хиральной чистотой, считается одной из фундаментальнейших характеристик живого. А поскольку при любом абиогенном синтезе (например, в аппарате Миллера) образующиеся аминокислоты будут состоять из приблизительно равных (по теории вероятностей) долей право– и левовращающих изомеров, то в дальнейшем – при синтезе из этого «сырья» белков – перед нами встанет задача: как химическими методами разделить смесь веществ, которые по определению химически идентичны? (Не зря оптической активностью обладают лишь природные сахара – и ни один из синтетических, а упомянутые выше полипептиды из метеоритного вещества состоят из равных долей право– и левовращающих аминокислот.)
Между тем, даже успешный синтез «живых» макромолекул (до которого еще, что называется, «семь верст – и все лесом») сам по себе проблемы не решает. Для того, чтобы макромолекулы заработали, они должны быть организованы в клетку – причем никаких возможностей для «промежуточной посадки» в ходе этого немыслимой сложности «перелета» вроде бы не просматривается: все так называемые доклеточные формы жизни – вирусы – являются облигатными (т.е. обязательными) внутриклеточными паразитами, а потому навряд ли могут являться предшествениками клеток. Пропасть, отделяющая полный набор аминокислот и нуклеотидов от простейшей по устройству бактериальной клетки, в свете современных знаний стала казаться еще более непреодолимой, чем это представлялось в прошлом веке.
Известна такая аналогия: вероятность случайного возникновения осмысленной аминокислотно-нуклеотидной последовательности соответствует вероятности того, что несколько килограммов типографского шрифта, будучи сброшены с крыши небоскреба, сложатся в 105-ую страницу романа «Война и мир». Абиогенез (в его классическом виде) как раз и предполагал такое «сбрасывание шрифта» – раз, 10 раз, 10100 раз – сколько понадобится, пока тот не сложится в требуемую страницу. Сейчас всем понятно, что это просто несерьезно: потребное для этого время (его вполне можно рассчитать) на много порядков превосходит время существования всей нашей Вселенной (не более 20 млрд лет). В результате мы оказываемся перед неизбежной необходимостью признать прямое вмешательство в этот случайный процесс Бога (тут можно придумать какие-нибудь эвфемизмы, но суть от этого не изменится); а раз так, то данная проблема, как легко догадаться, вообще не относится к сфере науки. Таким образом, получается, что по крайней мере в рамках чисто химического подхода проблема зарождения жизни принципиально неразрешима.
В качестве альтернативы абиогенезу выступала концепция панспермии, связанная с именами таких выдающихся ученых, как Г. Гельмгольц, У. Томпсон (лорд Кельвин), С. Аррениус, В.И. Вернадский. Эти исследователи полагали, что жизнь столь же вечна и повсеместна, как материя, и зародыши ее постоянно путешествуют по космосу; Аррениус, в частности, доказал путем расчетов принципиальную возможность переноса бактериальных спор с планеты на планету под действием давления света; предполагалось также, что вещество Земли в момент ее образования из газо-пылевого облака уже было «инфицировано» входившими в состав последнего «зародышами жизни».
Концепцию панспермии обычно упрекают в том, что она не дает принципиального ответа на вопрос о путях происхождения жизни, и лишь отодвигает решение этой проблемы на неопределенный срок. При этом молчаливо подразумевается, что жизнь должна была произойти в некой конкретной точке (или нескольких точках) Вселенной, и далее расселяться по космическому пространству – подобно тому, как вновь возникшие виды животных и растений расселяются по Земле из района своего происхождения; в такой интерпретации гипотеза панспермии действительно выглядит просто уходом от решения поставленной задачи. Однако действительная суть этой концепции заключается вовсе не в романтических межпланетных странствиях «зародышей жизни», а в том, что жизнь как таковая просто является одним из фундаментальных свойств материи, и вопрос о «происхождении жизни» стоит в том же ряду, что и, например, вопрос о «происхождении гравитации».
Легко видеть, что из двух исходных положений концепции панспермии – вечность жизни и повсеместность ее распространения – фальсифицируемым (т.е. проверяемым – см. дополнительную главу 1-а) является лишь второе. Однако все попытки обнаружить живые существа (или их ископаемые остатки) вне Земли, и прежде всего – в составе метеоритного вещества, так и не дали положительного результата. Неоднократно появлявшиеся сообщения о находках следов жизни на метеоритах основаны или на ошибочной интерпретации некоторых бактериоподобных неорганических включений, или на загрязнении «небесных камней» земными микроорганизмами. Метеоритное вещество оказалось достаточно богатым органикой, однако вся она, как уже было сказано, не обладает хиральной чистотой; это последнее обстоятельство – весьма сильный довод против принципиальной возможности существования «межзвездной жизни». Таким образом, по крайней мере положение, касающееся повсеместности распространения жизни во Вселенной, не нашло подтверждения. Это заставляет сделать грустный вывод, что панспермия, так же как и абиогенез, не дает удовлетворительного ответа на вопрос о возникновении жизни на Земле
.
Реальный прорыв в этой области обозначился лишь в последние 20-25 лет, и связан он был с приложением к проблеме возникновения жизни теории самоорганизующихся систем. Самоорганизующейся называют такую систему, которая обладает способностью корректировать свое поведение на основе предшествующего опыта (сам термин был введен в 1947 г. одним из создателей кибернетики физиологом У. Эшби). Следует сразу оговорить, что при этом было строго показано, что рассмотрение процессов развития (в том числе – биологических систем из добиологических) принципиально невозможно в рамках классической термодинамики. Создатель альтернативной, неравновесной, термодинамики И. Пригожин произвел научную революцию тех же примерно масштабов, что в свое время Ньютон или Эйнштейн, и революция эта еще отнюдь не завершена (подробнее обо всем этом – см. дополнительную главу 4-а). Между тем, все красивые модели последних лет – разрушение зеркальной симметрии с возникновением хиральной чистоты В.И. Гольданского (1986), или более подробно рассматриваемые далее гиперциклы М. Эйгена (1982) – работают только в рамках пригожинской термодинамики. Именно поэтому все они не имеют отношения к классическому абиогенезу: если Геккель и Опарин сводили биологию к химии, то физхимик Эйген, как мы увидим, в известном смысле сводит химию к биологии.
Эйген выдвинул концепцию образования упорядоченных макромолекул из неупорядоченного вещества на основе матричной репродукци и естественного отбора. Он начинает с того, что дарвиновский принцип естественного отбора (ЕО) – единственный понятный нам способ создания новой информации (это физическая величина, отражающая меру упорядоченности системы). Если имеется система самовоспроизводящихся единиц, которые строятся из материала, поступающего в ограниченном количестве из единого источника, то в ней с неизбежностью возникает конкуренция и, как ее следствие, ЕО. Эволюционное поведение, управляемое ЕО, основано на самовоспроизведении с «информационным шумом» (в случае эволюции биологических видов роль «шума» выполняют мутации). Наличия этих двух физических свойств достаточно, чтобы стало принципиально возможным возникновение системы с прогрессирующей степенью сложности.
В этом плане предшественником Эйгена является биохимик Г.Кастлер (1966), проанализировавший поведение системы нуклеиновых кислот в рамках теории информации. Он пришел к выводу, что новая информация возникает в системе, только если в ней происходит случайный выбор («методом тыка») с последующим запоминанием его результатов, а не целенаправленный отбор наилучшего варианта. В последнем случае можно говорить лишь о реализации той информации, что заложена в систему изначально, то есть о выделении уже имеющейся информации из «шума». Сама же возможность возникновения «новизны» (т.е. акта творчества) определяется свойствами информации как таковой: как было показано А.А. Ляпуновым (1965), на нее не распространяются законы сохранения, т.е. информация, в отличие от материи и энергии, может быть заново создана (и, соответственно, может быть и безвозвратно утрачена).
Говоря об усложнении системы, необходимо упомянуть выводы еще одного основоположника кибернетики, Дж. фон Неймана (1960), решавшего проблему самовоспроизведения автоматов. Оказалось, способность к самовоспроизведению принципиально зависит от сложности организации. На низшем уровне сложность является вырождающейся, т.е. каждый автомат способен воспроизводит лишь менее сложные автоматы. Существует, однако, вполне определенный критический уровень сложности, начиная с которого эта склонность к вырождению перестает быть всеобщей: «Сложность, точно так же, как и структура организмов, ниже некого минимального уровня является вырождающейся, а выше этого уровня становится самоподдерживающейся или даже может расти».
Итак, Эйгену «всего-навсего» осталось найти реальный класс химических реакций, компоненты которых вели бы себя подобно дарвиновским видам, т.е. обладали бы способностью «отбираться» и, соответственно, эволюционировать в сторону увеличения сложности организации. Именно такими свойствами, как выяснилось, и обладают нелинейные автокаталитические цепи, названные им гиперциклами. Здесь необходимо дать некоторые пояснения.
Простейшим случаем каталитической реакции является превращение исходного вещества (субстрат – S) в конечное (продукт – P) при участии единственного фермента (E); уже этот механизм требует по меньшей мере трехчленного цикла, который называется реакционным (рисунок 14, а). Существуют, однако, и гораздо более сложные реакционные циклы. Таков, например, цикл Кребса – 12-членный цикл, лежащий в основе клеточного дыхания: он катализирует превращение молекулы двухатомной уксусной кислоты (в форме ацетил-кофермента a – CH3CO Koa) в 2 молекулы CO2 и 8 атомов H (рисунок 14, б). Другой пример – углеродный цикл Бете-Вайцзекера, обеспечивающий светимость Солнца за счет превращения четырех атомов водорода 1H в атом гелия 4He (рисунок 14, в). Несмотря на серьезнейшие различия между этими реакциями (первая является химической, а вторая – ядерной), они обладают фундаментальным сходством: в обеих высокоэнергетическое вещество превращается в продукты, бедные энергией, при сохранении – т.е. циклическом воспроизведении – промежуточных компонентов (интермедиатов).
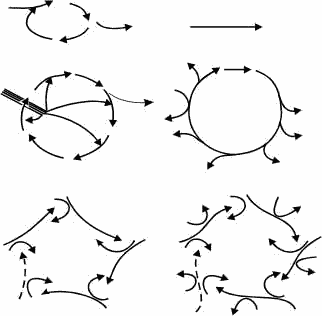
Следующий за реакционным циклом уровень организации представляет собой каталитический цикл, в котором некоторые – или все – интермедиаты сами являются катализаторами для одной из последующих реакций. Каждый из них (Ei+1) образуется из высокоэнергетического субстрата (S) при каталитической поддержке от предыдущего интермедиата (Ei) (рисунок 14, г). Таким образом, каталитический цикл как целое эквивалентен автокатализатору.
Если же такие автокаталитические (т.е. самовоспроизводящиеся) единицы оказываются, в свою очередь, сочленены между собой посредством циклической связи, то возникает каталитический гиперцикл. Гиперцикл, таким образом, основан на нелинейном автокатализе – автокатализе как минимум второго порядка, и представляет собой следующий, более высокий уровень в иерархии автокаталитических систем. Он состоит из самоинструктирующихся единиц (Ii) с двойными каталитическими функциями: в качестве автокатализатора интермедиат Ii способен инструктировать свое собственное воспроизведение, и при этом катализирует воспроизведение из высокоэнеогетического субстрата (S) следующего в цепи интермедиата (Ii+1) (рисунок 14, д).
Гиперциклы (одним из простейших примеров которых является размножение РНК-содержащего вируса в бактериальной клетке) обладают рядом уникальных свойств, порождающих дарвиновское поведение системы. Гиперцикл конкурирует (и даже более ожесточенно, чем дарвиновские виды) с любой самовоспроизводящейся единицей, не являющейся его членом; он не может стабильно сосуществовать и с другими гиперциклами – если только не объединен с ними в автокаталитический цикл следующего, более высокого, порядка. Состоя из самостоятельных самовоспроизводящихся единиц (что гарантирует сохранение фиксированного количества информации, передающейся от «предков» к «потомкам»), он обладает и интегрирующими свойствами. Таким образом, гиперцикл объединяет эти единицы в систему, способную к согласованной эволюции, где преимущества одного индивида могут использоваться всеми ее членами, причем система как целое продолжает интенсивно конкурировать с любой единицей иного состава.
Итак, именно гиперцикл (который сам по себе – еще чистая химия) является тем самым критическим уровнем, начиная с которого сложность неймановского «самовоспроизводящегося автомата» перестает быть вырождающейся. Эта концепция, в частности, вполне удовлетворительно описывает возникновение на основе взаимного катализа системы «нуклеиновая кислота-белок» – решающее событие в процессе возникновения жизни на Земле. Вместе с тем, сам Эйген подчеркивает, что в ходе реальной эволюции гиперцикл вполне мог «вымереть» – после того, как ферментные системы следующего поколения (с более высокой точностью репродукции) сумели индивидуализировать интегральную систему в форме клетки.
Однако на процесс возникновения жизни можно посмотреть и с несколько иной позиции, не биохимической , а геохимической, как это делает, например, А.С. Раутиан (1995). Мы уже говорили о том, что с общепланетарной точки зрения жизнь – это способ упорядочения и стабилизации геохимических круговоротов; откуда же берется сам геохимический круговорот?
Открытый космос холоден (лишь на 4о теплее абсолютного нуля) потому, что концентрация вещества в нем ничтожно мала (3*10-31 г/см3), и звездам просто нечего нагревать; по этой же самой причине, кстати сказать, Вселенная прозрачна, и мы видим небесные светила. В то же время любая планета, будучи непрозрачной, аккумулирует часть энергии, излучаемой центральным светилом и нагревается, и тогда между нагретой планетой и холодным космосом возникает температурный градиент ТГ. Если планета обладает при этом достаточно подвижной газообразной и/или жидкой оболочкой (атмосферой и/или гидросферой), то ТГ с неизбежностью порождает в ней – просто за счет конвекции – физико-химический круговорот. В этот круговорот с неизбежностью же вовлекается и твердая оболочка планеты (в случае Земли – кора выветривания), в результате чего возникает глобальный геохимический цикл – прообраз биосферы.
Конец бесплатного ознакомительного фрагмента.