Неоднозначная зависимость величин наблюдается в любых процессах, т.к. для изменения состояния тела всегда требуется определённое время (время
) и реакция тела отстаёт от вызывающих её причин. Такое отставание тем меньше, чем медленнее изменяются внешние условия Однако для некоторых процессов отставание при замедлении изменения внешних условий не уменьшается. В этих случаях неоднозначную зависимость величин называется гистерезисной, а само явление — Г.
Г. наблюдается в различных веществах и при разных физических процессах. Наибольший интерес представляют: магнитный Г., диэлектрический Г. и упругий Г.
Магнитный Г. наблюдается в магнитных материалах, например в
. Основной особенностью ферромагнетиков является наличие спонтанной (самопроизвольной) намагниченности. Обычно ферромагнетик намагничен не однородно, а разбит на доме'ны — области однородной спонтанной намагниченности, у которых величина намагниченности (магнитного момента единицы объема) одинакова, а направления различны. Под действием внешнего магнитного поля число и размеры доменов, намагниченных по полю, увеличиваются за счёт др. доменов. Кроме того, магнитные моменты отдельных доменов могут поворачиваться по полю. В результате магнитный момент образца увеличивается.
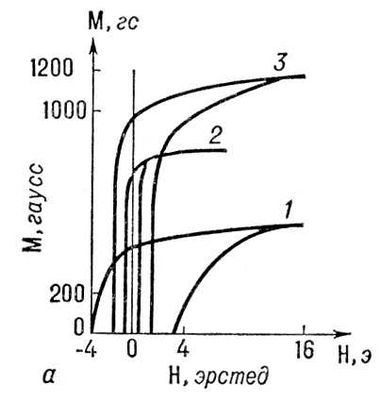
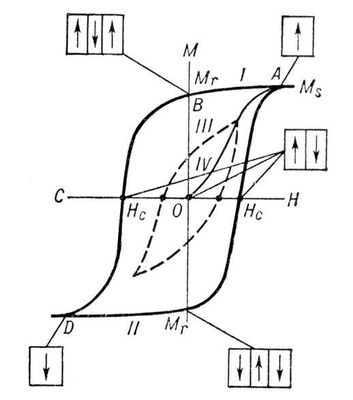
На
рис. 1
изображена зависимость магнитного момента
Мферромагнитного образца от напряжённости
Нвнешнего магнитного поля (кривая намагничивания). В достаточно сильном магнитном поле образец намагничивается до насыщения (при дальнейшем увеличении поля значение М практически не изменяется, точка А). При этом образец состоит из одного домена с магнитным моментом насыщения
M
s, направленным по полю. При уменьшении напряжённости внешнего магнитного поля
Нмагнитный момент образца
Мбудет уменьшаться по кривой
Iпреимущественно за счёт возникновения и роста доменов с магнитным моментом, направленным против поля. Рост доменов обусловлен движением доменных стенок. Это движение затруднено из-за наличия в образце различных дефектов (примесей, неоднородностей и т.п.), которые закрепляют доменные стенки в некоторых положениях; требуются достаточно сильные магнитные поля для того, чтобы их сдвинуть. Поэтому при уменьшении поля
Ндо нуля у образца сохраняется т. н. остаточный магнитный момент
M
r(точка
В).
Образец полностью размагничивается лишь в достаточно сильном поле противоположного направления, называемом коэрцитивным полем (
)
Н
с(точка С). При дальнейшем увеличении магнитного поля обратного направления образец вновь намагничивается вдоль поля до насыщения (точка
D). Перемагничивание образца (из точки
Dв точку
А) происходит по кривой
II. Т. о., при циклическом изменении поля кривая, характеризующая изменение магнитного момента образца, образует петлю магнитного Г. Если поле
Нциклически изменять в таких пределах, что намагниченность насыщения не достигается, то получается непредельная петля магнитного Г. (кривая
III). Уменьшая амплитуду изменения поля
Ндо нуля, можно образец полностью размагнитить (прийти в точку О). Намагничивание образца из точки О происходит по кривой
IV.
При магнитном Г. одному и тому же значению напряжённости внешнего магнитного поля
Нсоответствуют разные значения магнитного момента
М. Эта неоднозначность обусловлена влиянием состояний образца, предшествующих данному (т. е. магнитной предысторией образца).
Вид и размеры петли магнитного Г., величина
Н
св различных ферромагнетиках могут меняться в широких пределах. Например, в чистом железе
Н
с=1
э, в сплаве магнико
Н
с=580 э. На петлю магнитного Г. сильно влияет обработка материала, при которой изменяется число дефектов (
рис. 2
).
Площадь петли магнитного Г. равна энергии, теряемой в образце за один цикл изменения поля. Эта энергия идёт, в конечном счёте, на нагревание образца. Такие потери энергии называются гистерезисными. В тех случаях, когда потери на Г. нежелательны (например, в сердечниках трансформаторов, в статорах и роторах электрических машин), применяют магнитномягкие материалы, обладающие малым
Н
си малой площадью петли Г. Для изготовления постоянных магнитов, напротив, требуются магнитножёсткие материалы с большим
Н
с.
С ростом частоты переменного магнитного поля (числа циклов перемагничивания в единицу времени) к гистерезисным потерям добавляются др. потери, связанные с
и
. Соответственно площадь петли Г. при высоких частотах увеличивается. Такую петлю иногда называют динамической петлей, в отличие от описанной выше статической петли.
От магнитного момента зависят многие др. свойства ферромагнетика, например электрическое сопротивление, механическая деформация. Изменение магнитного момента вызывает изменение и этих свойств. Соответственно наблюдается, например, гальваномагнитный Г., магнитострикционный Г.
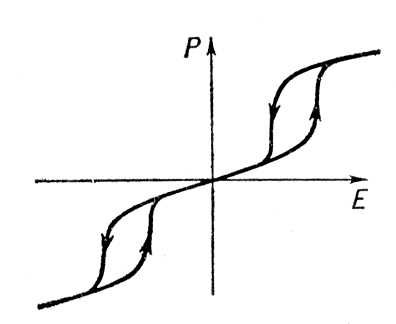
Диэлектрический Г. наблюдается обычно в
, например титанате бария. Зависимость поляризации
Рот напряжённости электрического поля
Ев сегнетоэлектриках (
рис. 3
) подобна зависимости
Мот
Нв ферромагнетиках и объясняется наличием спонтанной электрической поляризации, электрических
и трудностью перестройки доменной структуры. Гистерезисные потери составляют большую часть
в сегнетоэлектриках.
Поскольку с поляризацией связаны др. характеристики сегнетоэлектриков, например деформация, то с диэлектрическим Г. связаны др. виды Г., например пьезоэлектрический Г. (
рис. 4
), Г.
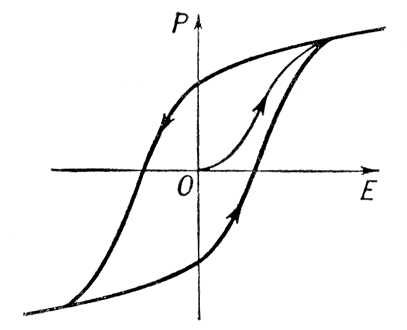
. В некоторых случаях наблюдаются двойные петли диэлектрического Г. (
рис. 5
). Это объясняется тем, что под влиянием электрического поля в образце происходит фазовый переход с перестройкой кристаллической структуры. Такого рода диэлектрический Г. тесно связан с Г. при фазовых переходах.
Упругий Г., т. е. гистерезисная зависимость деформации
иот механического напряжения
s, наблюдается в любых реальных материалах при достаточно больших напряжениях (
рис. 6
). Упругий Г. возникает всякий раз, когда имеет место пластическая (неупругая) деформация (см.
). Пластическая деформация обусловлена перемещением дефектов, например
, всегда присутствующих в реальных материалах. Примеси, включения и др. дефекты, а также сама кристаллическая решётка стремятся удержать дислокацию в определенных положениях в кристалле. Поэтому требуются напряжения достаточной величины, чтобы сдвинуть дислокацию. Механическая обработка и введение примесей приводят к закреплению дислокаций, в результате чего происходит упрочнение материала, пластическая деформация и упругий Г. наблюдаются при больших напряжениях. Энергия, теряемая в образце за один цикл, идёт в конечном счёте на нагревание образца. Потери на упругий Г. дают вклад во
. В случае упругих деформаций, помимо гистерезисных, есть и др. потери, например обусловленные
. Величина этих потерь, в отличие от гистерезисных, зависит от частоты изменения
s(или
и). Иногда понятие «упругий Г.» употребляется шире — говорят о динамической петле упругого Г., включающей все потери на данной частоте.
Лит.:Киренский Л. В., Магнетизм, 2 изд., М., 1967; Вонсовский С. В., Современное учение о магнетизме, М. — Л., 1952; Бозорт Р., Ферромагнетизм, пер. с англ., М., 1956; Иона Ф., Ширане Д., Сегнетоэлектрические кристаллы, пер. с англ., М., 1965; Постников В. С., Внутреннее трение в металлах, М., 1969; Физический энциклопедический словарь, т. 1, М., 1960.
А. П. Леванюк, Д. Г. Санников.
Рис. 2. Влияние механической и термической обработки на форму петли магнитного гистерезиса пермалоя: 1 — после наклёпа; 2 — после отжига; 3 — кривая мягкого железа (для сравнения).
Рис. 5. Двойная петля диэлектрического гистерезиса.
Рис. 6. Петля упругого гистерезиса: s — механическое напряжение; u — деформация.
Рис. 1. Петля магнитного гистерезиса для ферромагнетика: Н — напряжённость магнитного поля; М — магнитный момент образца; Н
с— коэрцитивное поле; M
r— остаточный магнитный момент; M
s— магнитный момент насыщения. Пунктиром показана непредельная петля гистерезиса. Схематически приведена доме'нная структура образца для некоторых точек петли.
Рис. 3. Петля диэлектрического гистерезиса в сегнетоэлектрике: Р — поляризация образца; Е — напряжённость электрического поля.
Рис. 4. Петля гистерезиса обратного пьезоэлектрического эффекта в титанате бария: U — деформация: Е — напряжённость электрического поля.
Гистерезисный электродвигатель
Гистере'зисный электродви'гатель,
, у которого вращающий момент возникает за счёт гистерезиса при перемагничивании массивного ротора с сердечником из магнитного материала, имеющего широкую петлю гистерезиса. При мощностях до 100
вти частоте 400
гц. Г. э. обладают несколько лучшими по сравнению с синхронными электродвигателями энергетическими характеристиками. Г. э. надёжны в эксплуатации и долговечны, они бесшумны и способны работать с различной частотой вращения. Г. э. широко применяются в электроприводе малой мощности и в системах автоматизированного управления. В автоматических приборах управления применяются реактивно-гистерезисные синхронные двигатели мощностью 10—15
мквтс частотой вращения, не превышающей несколько
об/мин, и кпд менее 1%.
Лит.:Бертинов А. И., Ермилов М. А., Гистерезисные электродвигатели, М., 1967; Арменский Е. В., Фалк Г. Б., Электрические микромашины, М., 1968.
В. А. Прокудин.
Гистероскоп
Гистероско'п(от греч. hystera — матка и
), прибор для осмотра внутренней полости матки; один из приборов для
. Состоит из металлической трубки и оптического аппарата, представляющего собой систему призм и нескольких линз и снабженного на конце электрической лампочкой. Фотоприставка к прибору позволяет фотографировать внутреннюю поверхность матки.
Гистидин
Гистиди'н, a-амино-b-имидазолилпропионовая кислота:

, обладающая основными свойствами, незаменимая для многих животных; организм человека способен к ограниченному синтезу Г. Входит в состав активных центров многих
, в частности рибонуклеазы, транскетолазы. Начальная стадия ферментативного разрушения Г. в организме — отщепление аммиака с образованием уроканиновой кислоты, выводящейся с мочой. Реакция
Г. необратима, катализирует её фермент гистидин-аммиак-лиаза (гистидин-a-дезаминаза), обнаруженный в печени животных и у бактерий. Недостаток Г. приводит ко многим нарушениям обмена веществ, в том числе к торможению синтеза
. Г. — предшественник специфических
скелетной мускулатуры —
и
. Декарбоксилирование Г. ведёт к образованию биологически активного амина —
; этот процесс катализирует гистидин-декарбоксилаза — фермент, относящийся к классу
. Фермент действует только на L-изомер (природную форму) Г. Реакция обратимо тормозится
дыхания — цианидом, гидроксиламином, семикарбазидом.
А. А. Болдырев, Е. В. Петушкова.
Гистиоциты
Гистиоци'ты(от греч. histion — ткань и kytos — вместилище, здесь — клетка), блуждающие клетки в покое, полибласты, клазматоциты, клетки рыхлой соединительной ткани у позвоночных животных и человека. Резко контурированы, с базофильной цитоплазмой, в которой часто встречаются вакуоли и включения, форма клетки варьирует в связи с её способностью к амёбоидному движению. Г. выполняют защитную функцию, захватывая и переваривая различные посторонние частички (в т. ч. и бактерии). При различного рода раздражениях, например при воспалительных реакциях, Г. активизируются, превращаясь в типичные
. Иногда цитоплазма Г. образует короткие закруглённые отростки, отрывающиеся от тела клетки (клазматоз). У зародышей Г. развиваются из
, во взрослом организме — из недифференцированных клеток рыхлой соединительной ткани, ретикулярной ткани и некоторых видов кровяных клеток —
и
.
Е. С. Кирпичникова.
Гисто-гематические барьеры
Ги'сто-гемати'ческие барье'ры, гемато-паренхиматозные, тканевые, гистиоцитарные барьеры, механизмы, регулирующие обмен между общей внутренней средой организма — кровью и непосредственно питательной средой органов и тканей — тканевой, или внеклеточной, жидкостью. Анатомическая основа Г.-г. б. — эндотелий капилляров и прекапилляров. Термин «Г.-г. б.» введён сов. физиологом Л. С. Штерн (1929). Г.-г. б. выполняют также защитную функцию, препятствуя переходу из крови в ткани и из тканей в кровь вредных и чужеродных веществ. Этим объясняется как неравномерное распределение многих веществ в организме, так и отсутствие эффекта при лечении некоторыми лекарственными препаратами. Приспособляемость Г.-г. б. к условиям внешней и внутренней среды является одним из важнейших условий поддержания постоянства внутренней среды (
), устойчивости физиологических функций, предохранения от инфекций, интоксикаций и т.п. См. также
,
.
Лит.:Штерн Л. С., Непосредственная питательная среда органов и тканей. Физиологические механизмы, определяющие ее состав и свойства. Избранные труды, М., 1960; Физиология и патология гисто-гематических барьеров, М., 1968.
Г. Н. Кассиль.
Гистогенез
Гистогене'з(от греч. histos — ткань и
), развитие тканей, совокупность закономерно протекающих процессов, обеспечивающих возникновение, существование и восстановление тканей животных организмов с их специфическими в разных органах свойствами. Изучение Г. разных тканей и его закономерностей — одна из важнейших задач
. Термином «Г.» принято обозначать развитие тканей в
. Однако закономерности Г. не могут рассматриваться в отрыве от эволюционного развития тканей (филогистогенеза). В основе Г. лежит начинающаяся с самых ранних стадий
клеточная
— развитие нарастающих морфо-функциональных различий между специализирующимися клетками. Это сложный молекулярно-генетический процесс закономерного включения активности
, определяющих специфику белковых синтезов в клетке. Размножение клеток, их взаимоперемещения и др. процессы приводят к формированию эмбриональных зачатков, представляющих собой группы клеток, закономерно расположенные в теле зародыша. В результате тканевой дифференцировки эмбриональных зачатков возникает всё многообразие тканей разных органов тела. В послезародышевом периоде процессы Г. подразделяют на 3 основных типа: в тканях, клетки которых не размножаются (например, нервная ткань); в тканях, размножение клеток которых связано главным образом с ростом органа (например, паренхима пищеварительных. желёз, почек); в тканях, характеризующихся постоянным обновлением клеток (например, кроветворная ткань, многие покровные эпителии). Совокупность клеток, совершающих определенный Г., подразделяют на ряд последовательных групп (фондов): фонд родоначальных клеток, способных как к дифференцировке, так и к восполнению убыли себе подобных; фонд клеток-предшественников, дифференцирующихся и способных к размножению; фонд зрелых, закончивших дифференцировку клеток. Восстановление поврежденных или частично утраченных тканей после травм осуществляется благодаря т. н. репаративному Г. При патологических условиях процессы Г. могут подвергнуться глубоким качественным изменениям и привести к развитию опухолевых тканей (см.
).
Лит.:Хлопин Н. Г., Общебиологические и экспериментальные основы гистологии, М., 1946 (библ.); Заварзин А. А., Очерки эволюционной гистологии крови и соединительной ткани, Избр. труды, т. 4, М. — Л., 1953; Хрущов Н. Г., Функциональная цитохимия рыхлой соединительной ткани, М., 1969 (библ.).
Н. Г. Хрущов.
Гистограмма
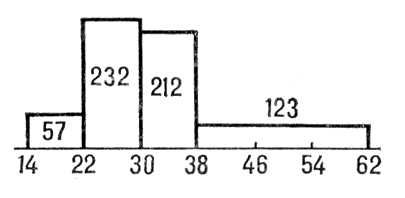
Гистогра'мма(от греч. histos, здесь — столб и
), столбчатая диаграмма, один из видов графического изображения статистического распределении каких-либо величин по количественному признаку. Г. представляет собой совокупность смежных прямоугольников, построенных на прямой линии. Площадь каждого прямоугольника пропорциональна частоте нахождения данной величины в изучаемой совокупности. Пусть, например, измерение диаметров стволов 624 сосен дало следующие результаты:
Диаметр,
см
14—22
22—30
30—38
38—62
Число стволов
57
232
212
123
На горизонтальной оси откладываются границы групп, на которые стволы разбиты по их диаметру, и на отрезке, соответствующем каждой группе, строится как на основании прямоугольник с площадью, пропорциональной числу стволов, попавших в данную группу (
рис. 1
).
В виде Г. часто изображают гранулометрический состав горных пород. В этом случае на вертикальной оси откладывают процентное содержание полученных групп частиц т. н. фракций, а на горизонтальной оси — логарифмы их граничных размеров (
рис. 2
). Использование логарифмов вызвано тем, что при гранулометрическом анализе частицы подразделяются на фракции, размеры которых убывают в геометрической прогрессии. Иногда Г. строятся на произвольно выбранных равных отрезках, независимо от разности граничных размеров фракций. Тогда высоты столбиков пропорциональны содержанию размеров фракций.
Рис. 1 к ст. Гистограмма.
Рис. 2 к ст. Гистограмма.
Гистология
Гистоло'гия(от греч. histos — ткань и
), наука о
многоклеточных животных и человека. Изучением тканей растений занимается
. Название «Г.» введено немецким учёным К. Майером (1819). Задачи Г. — выяснение эволюции тканей, исследование их развития в организме (
), строения и функции специализированных клеток, межуточных сред, взаимодействия клеток в пределах одной ткани и между клетками разных тканей, регенерации тканевых структур и регуляторных механизмов, обеспечивающих целостность и совместную деятельность тканей. Основной предмет изучения Г. — комплексы клеток в их взаимодействии друг с другом и с межуточными средами. Современная Г. уделяет много внимания изучению специфических особенностей клеток различных тканей; в этом разделе Г. и по методам исследования, и по технике имеет много общего с
, наукой об общих свойствах клеток. Г. принято разделять на общую Г., исследующую основные принципы развития, строения и функций тканей, и частную Г., выясняющую свойства тканевых комплексов в составе органов многоклеточных животных. Специальные разделы общей и частной Г. ставят своими задачами изучение химии тканей —
, и механизмов их деятельности — гистофизиология.
Исторический очерк.Становление Г. как самостоятельного раздела науки с 20-х гг. 19 в. связано с развитием
. Но ещё задолго до этого было отмечено, что органы животных состоят из компонентов, различающихся цветом и плотностью. По этим критериям Аристотель (4 в. до н. э.) выделял в составе органа «однородные части». Классификация «однородных частей» Аристотеля на протяжении столетий воспроизводилась в трудах учёных древности и средневековья вплоть до эпохи Возрождения. Сведения об «однородных частях» имеются в книгах римского врача и естествоиспытателя К. Галена (2 в. н. э.), среднеазиатского учёного Авиценны (10 в.) и итальянского врача и анатома Г. Фаллопия (16 в.). Изобретение в 17 в. микроскопа не сразу сказалось на уровне знаний о тонком строении органов. Первые микроскописты (англичане Р. Гук, Н. Грю, итальянец М. Мальпиги и голландец А. Левенгук) видели некоторые крупные клетки, кровеносные капилляры, нервы, но наблюдения эти были несистематичны и не связывались с анатомическими данными того времени. Даже к началу 19 в. представление о тканях основывалось, как и во времена Аристотеля, на оценке их невооружённым глазом. «Макроскопический» (домикроскопический) период развития Г. завершился фундаментальным трудом французского анатома и физиолога М. Биша «Общая анатомия в приложении к физиологии и медицине» (1802). Для обозначения частей органов Биша использовал термин «ткань», ранее предложенный Н. Грю в труде «Анатомия растений» (1672). При разграничении тканей Биша не только описывал компоненты разреза органа, но пытался выявить их свойства: отношение к разным реактивам, нагреванию и др. воздействиям. Биша различал 21 ткань. Предложенная им классификация была несовершенна, но сыграла прогрессивную роль в становлении Г. и позволила наряду с накоплением данных микроскопических исследований уже в 1-й четверти 19 в. сформулировать задачи Г. как самостоятельной науки. В 1819 вышла работа нем. учёного К. Майера «О гистологии и новом подразделении тканей человека», закрепившая понятие «ткань», В этой работе и особенно в монографии нем. учёного К. Гейзингера «Система гистологии» (1822) были сформулированы задачи Г., отличные от задач анатомии.
Интенсивное развитие Г. началось с 30-х гг. 19 в. В эти и последующие годы был существенно усовершенствован микроскоп. Развивалась и техника подготовки тканей для микроскопии. Методологической основой Г. становится
, окончательно обоснованная нем. биологом Т. Шванном в 1839. В 1-й половине 19 в. большое количество данных о микроскопическом строении органов и тканей было получено чешским учёным Я. Пуркине, немецкими учёными И. Мюллером, Я. Генле, Т. Шванном, Р. Ремаком и русскими — Н. М. Якубовичем, Н. Ф. Овсянниковым, Обобщение обширной литературы и собственные исследования позволили немецким гистологам Ф. Лейдигу (1853) и А. Кёлликеру (1855) создать рациональную классификацию тканей, сохранившуюся в общих чертах до настоящего времени. В системах Лейдига и Кёлликера выделялись 4 группы тканей не только по структуре, но и по функциональному значению в организме: эпителиальная, соединительная, мышечная и нервная. Последующее углубление морфо-физиологической классификации Лейдига и Кёлликера (главным образом при изучении развития тканей) заложило основы современной Г.
Во 2-й половине 19 — начале 20 вв. были получены существенные данные об эпителиальных тканях (А. Кёлликером, франц. учёными Э. Лагесом, Л. Ранвье и русским учёным С. Г. Часовниковым), о тканях русскими учёными И. И. Мечниковым, Ф. Гойером, В. Данчаковой и особенно А. А. Максимовым, создавшим и детально обосновавшим оригинальную теорию гистогенетического единства тканей внутренней среды, получившую впоследствии, в частности в 50—60-е гг. 20 в., экспериментальные подтверждения), о мышечных тканях (немецким гистологом М. Гейденгайном, русским биологом А. И. Бабухиным, Л. Ранвье), о нервной ткани (итальянским гистологом К. Гольджи, русскими — М. Д. Лавдовским, В. Я. Рубашкиным, А. С. Догелем, испанским — С. Рамон-и-Кахалем). К этому времени относятся крупные открытия в области общей цитологии: описание непрямого деления ядра и клетки —
(русские учёные А. Шнейдер, И. Д. Чистяков, немецкие — В. Флемминг, Э. Страсбургер), открытие и изучение цитоплазматических органоидов —
,
(немецкие учёные Р. Альтман, К. Бенда, итальянский — К. Гольджи). Открытие И. И. Мечниковым клеточной природы воспалительного процесса сблизило цитологию и Г. с проблемами патологии. Этому в большой мере способствовали труды немецкого учёного Р. Вирхова. Г. всё более сближалась с физиологией, что прослеживается в трудах французских учёных О. Пренана, А. Поликара, немецких — О. Гертвига, М. Гейденгайна, русского учёного И. Ф. Огнева. Большое значение для развития Г. и цитологии имела книга О. Гертвига «Клетки и ткани» (1893—98), в которой были обобщены многочисленные микроскопические исследования и сделан вывод, что углубленное изучение клетки — путь решения многих биологических проблем, в том числе и выяснения тканевых взаимоотношений.
В России Г. развивалась в Петербургском (Н. М. Якубович, М. Д. Лавдовский, А. С. Догель), Московском (А. И. Бабухин, И. Ф. Огнев, В. П. Карпов), Казанском (Н. Ф. Овсянников, К. А. Арнштейн, А. Н. Миславский), Киевском (М. И. Перемежко) университетах. После Октябрьской революции, кроме кафедр университетов, Г. начала разрабатываться и в медицинских институтах, где сложились школы А. А. Заварзина, Н. Г. Хлопина, Б. И. Лаврентьева, М. А. Барона. Гистологические исследования проводятся также в институтах и в лабораториях АН СССР и АМН СССР. Советские гистологи внесли большой вклад в познание свойств тканей, вскрыли многие важные закономерности в гистогенезах и особенностях функционирования тканевых структур. Существенно усовершенствованы гистохимические методы исследования, с помощью которых получены данные о развитии, функционировании и патологии тканей.
Предмет, задачи и методы Г.Историческое развитие многоклеточных животных (
) привело к дифференцированию и специализации клеток и обособлению клеточных систем и комплексов, выполняющих определенные функции. Тканями принято считать филогенетически сложившиеся системы клеток, объединённые общей структурой, функцией и происхождением. По этим признакам выделяют: эпителии, образующие внешние или внутренние покровы организма и различные железы, выполняющие защитную, пищеварительную и эндокринную функции; ткани внутренней среды (соединительная ткань, кровь), принимающие основное участие во внутреннем трофике и несущие опорные функции; мышечную ткань, выполняющую сократительную функции; нервную ткань, осуществляющую основную регуляцию жизнедеятельности всех систем организма. В любом органе многоклеточных животных сосуществуют и тесно взаимодействуют многочисленные разные ткани.
В современной Г., особенно в гистофизиологии, широко используют экспериментальные подходы к изучению свойств тканей. Из них часто применяют воспроизведение у подопытных животных процессов
,
, методику пересадок органов и их частей, экспериментальную денервацию тканей, стимуляцию и торможение деятельности тканей путём влияния на нервную и эндокринную системы или при помощи прямых влияний на отдельные синтезы, транспорт веществ, энергетику тканей и т.д. Для решения ряда задач Г. применяется метод тканевых и органных культур (см.
).
При изучении тканей широко используется цитологическая техника. Электронная микроскопия позволяет изучать субмикроскопическую структуру тканевых клеток, их морфологические контакты друг с другом и с межклеточными компонентами ткани. Гистохимия ставит своей задачей выяснение специфических особенностей обмена веществ в разных тканях. Преимущество этой методики перед биохимическим анализом — в возможности точной локализации тканевых процессов. Один из гистологических методов —
— позволяет исследовать кинетику клеточных популяций, гистогенезы, метаболическую активность тканей. Цитогенетический анализ, например при использовании хромосом-маркеров, применяется в опытах с
тканей.
Важная задача общей Г. — выяснение потенций развития, присущих каждому типу дифференцированных клеток, и механизмов, регулирующих сохранение постоянства дифференцировки и ее изменения. В каждой ткани различают несколько устойчивых типов клеточной дифференцировки, например фибробласты, образующие основное вещество соединительной ткани, и эритроидные клетки, образующие и несущие дыхательные пигменты. Каждый тип дифференцировки достигается в ходе многоэтапного процесса развития ткани — гистогенеза. В клетках, выполняющих специализированные функции, реализуется лишь небольшая часть возможностей, предусмотренных генетической программой организма. Остальная, не реализуемая в дифференцированных клетках часть
сохраняется в них, но находится в неактивном, или репрессированном, состоянии. При определенных внешних воздействиях на клетку может происходить дерепрессия, и характер дифференцировки клеток может изменяться. Такие изменения происходят во многих тканях постоянно, в частности при нормальном созревании входящих в их состав клеток, когда изменчивость клеток не выходит за типичные для каждой ткани пределы. В условиях же патологии наступают более значительные изменения дифференцировки тканевых клеток, называемые метаплазией.
Общая Г. исследует гистогенезы при формировании тканей в зародышевом развитии, а также при естественном обновлении тканей у взрослых животных, при регенерации после повреждений, вызвавших усиленную гибель клеток. С этим связана проблема детерминации клеток, участвующих в обновлении тканей, и факторов, регулирующих направление и темп процесса обновления. Клеточные популяции некоторых тканей, например нервной у взрослых животных, практически не обновляются. Нервные клетки обычно долго живут, но часть их всё же гибнет с возрастом в результате напряжений, заболеваний и т.д. В большинстве же тканей (эпителии и ткани внутренней среды) часть клеток сохраняет способность к делению. В таких тканях постоянно протекают процессы смены клеток. В нормальных условиях при обновлении клеточного состава гибель одних клеток компенсируется размножением других. Этот процесс обусловлен рядом регуляторных механизмов, действующих как внутри ткани, так и в организме в целом.
Длительное поддержание равновесного состояния в тканях, клетки которых имеют небольшой срок жизни (несколько дней или недель), обеспечивается особыми т. н. стволовыми клетками, способными к многократному делению. Стволовые клетки делятся и поддерживают собственную линию в организме в течение почти всей его жизни; они же дают начало развитию разных специализированных клеток данной ткани. Выяснение факторов, регулирующих размножение и дифференцировку стволовых клеток, а также механизмов, определяющих путь их развития, — важная проблема общей Г.