 |
|
Популярные авторы:: Борхес Хорхе Луис :: Азимов Айзек :: Грин Александр :: Толстой Лев Николаевич :: Чехов Антон Павлович :: Горький Максим :: Раззаков Федор :: Лавкрафт Говард Филлипс :: Лондон Джек :: Эллисон Харлан Популярные книги:: Справочник по реестру Windows XP :: Переселенцы Трансвааля :: Свидание :: Бык в загоне :: Облдрамтеатр :: Дуэль на Марсе :: Граф Монте-Кристо :: Здесь нет никого, кроме... :: Реанимация :: Дорога на Стрельну |
Большая Советская Энциклопедия (БИ)ModernLib.Net / Энциклопедии / БСЭ / Большая Советская Энциклопедия (БИ) - Чтение (стр. 27)
Количество периодических изданий, в которых публикуются работы по Б., очень велико и продолжает увеличиваться с каждым годом. Из зарубежных и международных журналов наиболее известны: «Journal of Biological Chemistry» (Balt., 1905-), «Biochemistry» (Wash., 1964-), «Archives of Biochemistry and Biophysics» (N. Y., 1942-), «Biochemical Journal» (L., 1906-), «Phytochemistry» (Oxf.- N. Y., 1962-), «Molecular Biology» (издаётся в Англии - журнал международный), «Bulletin de la Sociйtй de Chimie Biologique» (P., 1914-), «Enzymologia» (Haaga, 1936-), «Giornale di Biochimica» (Rome, 1955-),»Acta Biological et Medica Germanica»(Lpz., 1959-), «Hoppe Seyler's Zeitschrift fьr physiologische Chemie» (Berlin, 1877-), «Journal of Biochemistry». (Tokyo, 1922-). Популярны ежегодники: «Annual Review of Biochemistry» (Stan-ford, 1932-), «Advances in Enzymology and Related Subjects of Biochemistry» (N. Y., 1945-), «Advances in Protein Chemistry» (N. Y., 1945-), «Advances in Enzyme Regulation» (Oxf., 1963-), «Advances in Molecular Biology» и др. В СССР экспериментальные работы по Б. печатаются в журналах: «Биохимия» (М., 1936-), «Журнал эволюционной биохимии и физиологии» (М., 1965-), «Молекулярная биология» (М., 1967-), «Вопросы медицинской химии» (М., 1955-), «Украинский биохимический журнал» (К., 1926-), «Прикладная биохимия и микробиология» (М., 1965-), «Доклады АН СССР» (М., 1933-), «Бюллетень экспериментальной биологии и медицины» (М., 1936-), «Известия АН СССР. Серия биологии и медицины» (М., 1936-), «Известия АН СССР. Серия химическая» (М., 1936-), «Научные доклады высшей школы. Серия биологические науки» (М., 1958-) и в некоторых др. Обзорные работы по Б. печатаются в журнале «Успехи современной биологии» (М., 1932-), в ежегоднике «Успехи биологической химии» (т. 1-8, 1950-67), издаваемом Всесоюзным биохимическим обществом, в журнале «Успехи химии» (М., 1932-), «Реферативный журнал. Химия. Биологическая химия» (М., 1955-), в журнале Всесоюзного общества им. Менделеева. Часто выходят в свет труды биохимических институтов. Лит.:Руководства: Макеев И. А., Гулевич В. С., Броуде Л. М., Курс биологической химии, М., 1947; Кретович В. Л., Основы биохимии растений, 4 изд., М., 1964; 3барский Б. И., Иванов И. И., Мардашев С. Р., Биологическая химия, 4 изд., М., 1965; Фердман Д. Л., Биохимия, 3 изд., М., 1966. История: Прянишников Д., Избр. соч., т. 1, М., 1951, с. 5-19; Гулевич В. С., Избранные труды, М., 1954, с. 5-21; Парнас Я. О., Избранные труды, М., 1960, с. 5-10; Толкачевская Н. Ф., Развитие биохимии животных, М., 1963; Джуа М., История химии, пер. с итал., М., 1966; Развитие биологии в СССР, М., 1967; Кретович В.Л., Введение в энзимологию, М., 1967; Биохимия растений, пер. с англ., М., 1968; Lieben F., Geschichte der physiologischen Chemie, Lpz.-W., 1935. Монографии: Энгельгардт В. А., Некоторые проблемы современной биохимии, М., 1959; его же. Пути химии в познании явлений жизни, М., 1965; Северин С. Е., Биохимические основы жизни, М., 1961; Спирин А. С., Информационная РНК и биосинтез белков, М., 1962; Скулачев В. П., Соотношение окисления и фосфорилирования в дыхательной цепи, М., 1962; ферменты, под ред. А. Е. Браунштейна, М., 1964; Владимиров Г. Е., Пантелеева Н. С., Функциональная биохимия, Л., 1965; Ингрэм В., Биосинтез макромолекул, пер. с англ., М., 1966; Рэкер Э., Биоэнергетические механизмы, пер. с англ., М., 1967; Спирин А. С., Гаврилова Л. П., Рибосома, М., 1968. С. Е. Северин. «Биохимия» «Биохи'мия»,журнал, издаваемый АН СССР в Москве. Основан в 1936 А. Н. Бахом.Выходит 1 раз в 2 мес.В журнале публикуются экспериментальные работы по общей биохимии, энзимологии (учение о ферментах), обмену веществ в растениях, животных и микроорганизмах, по биохимии витаминов, гормонов и других природных физиологически активных соединений, а также по биохимическим основам технологии растительного и животного сырья. Тираж около 4 тыс. экз. (1970). О других биохимических журналах см. Биохимия . Биохор Биохо'р(от био... и греч. chMra - пространство), подразделение биосферы , представляющее собой группу сходных биотопов.Б. объединяются в биоциклы:суша, море и внутренние водоёмы. Биоценоз Биоцено'з(от био... и греч. koinуs - общий), совокупность растений, животных, микроорганизмов, населяющих участок суши или водоёма и характеризующихся определёнными отношениями как между собой, так и с абиотическими факторами среды (см. Биотоп ). Термин «Б.» был предложен немецким биологом К. Мебиусом (1877). Б. - комплекс организмов биогеоценоза , формирующийся в результате борьбы за существование, естественного отбора и других факторов эволюции. По участию в биогенном круговороте веществ в Б. различают три группы организмов. 1) Продуценты (производители) - автотрофные организмы , создающие органические вещества из неорганических; основные продуценты во всех Б. - зелёные растения (см. Фотосинтез ) .Деятельность продуцентов определяет исходное накопление органических веществ в Б. (см. Биомасса , Биологическая продуктивность ) .2) Консументы (потребители) - гетеротрофные организмы , питающиеся за счёт автотрофных. Консументы 1-го порядка - растительноядные животные, а также паразитические бактерии, грибы и другие бесхлорофильные растения, развивающиеся за счёт живых растений. Консументы 2-го порядка - хищники и паразиты растительноядных организмов. Бывают консументы 3-го и 4-го порядков (сверхпаразиты, суперпаразиты и т.п.), но всего в цепях питания не более 5 звеньев. На каждом последующем трофическом уровне количество биомассы резко снижается. Деятельность консументов способствует превращениям и перемещениям органических веществ в Б., частичной их минерализации, а также рассеянию энергии, накопленной продуцентами. 3) Редуценты (восстановители) - животные, питающиеся разлагающимися остатками организмов ( сапрофаги ), и особенно непаразитирующие гетеротрофные микроорганизмы - способствуют минерализации органических веществ, их переходу в усвояемое продуцентами состояние. Взаимосвязи организмов в Б. многообразны. Кроме трофических связей, определяющих цепи питания (иногда очень своеобразные - см. Паразитизм , Симбиоз ), существуют связи, основанные на том, что одни организмы становятся субстратом для других (топические связи), создают необходимый микроклимат и т.п. Часто можно проследить в Б. группы видов, связанные с определённым видом и целиком зависящие от последнего ( консорции ) . Для Б. характерно разделение на более мелкие подчинённые единицы - мероценозы, т.е. закономерно слагающиеся комплексы, зависящие от Б. в целом (например, комплекс обитателей гниющих дубовых пней в дубраве). Если энергетическим источником Б. служат не автотрофы, а животные (например, летучие мыши в Б. пещер), то такие Б. зависят от притока энергии извне и являются неполноценными, представляя в сущности мероценозы. В Б. можно выделить и другие подчинённые группировки организмов, например синузии.Для Б. также характерно разделение на группировки организмов по вертикали (ярусы Б.). В годовом цикле в Б. изменяются численность, стадии развития и активность отдельных видов, создаются закономерные сезонные аспекты Б. Б. - диалектически развивающееся единство, меняющееся в результате деятельности входящих в него компонентов, вследствие чего происходят закономерные изменение и смена Б. ( сукцессии ), которые могут приводить к восстановлению резко нарушенных Б. (например, леса после пожара и т.п.). Различают насыщенные и ненасыщенные Б. В насыщенном Б. все экологические ниши (см. Ниша экологическая ) заняты и вселение нового вида невозможно без уничтожения или последующего вытеснения какого-либо компонента Б. Ненасыщенные Б. характеризуются возможностью вселения в них новых видов без уничтожения других компонентов. Можно различать первичные Б., сложившиеся без воздействия человека (целинная степь, девственный лес), и вторичные, измененные деятельностью человека (леса, выросшие на месте сведённых, население водохранилищ). Особую категорию представляют агробиоценозы , где комплексы основных компонентов Б. сознательно регулируются человеком. Между первичными Б. и агробиоценозами имеется вся гамма переходов. Изучение Б. важно для рационального освоения земель и водных пространств, т.к. только правильное понимание регулятивных процессов в Б. позволяет человеку изымать часть продукции Б. без его нарушения и уничтожения. Лит.:Кашкаров Д. Н., Основы экологии животных, 2 изд., Л., 1945; Беклемишев В. Н., О классификации биоценологических (симфизиологических) связей, «Бюлл. Московского общества испытателей природы», 1951, т. 56, в. 5; Гиляров М. С., Вид, популяция и биоценоз, «Зоологический журнал», 1954, т. 33, в. 4; Арнольди К. В. и Л. В., О биоценозе, там же, 1963, т. 42, в. 2; Наумов Н. П., Экология животных, 2 изд., М., 1963; Основы лесной биогеоценологии, под ред. В. Н. Сукачева и Н. В. Дылиса, М., 1964; Макфедьен Э., Экология животных, пер. с англ., М., 1965; Одум Е., Экология, М., 1968; Дювиньо П. и Танг М., Биосфера и место в ней человека, пер. с франц., М., 1968; Tischler W., Synцkologie der Landtiere, Stuttg., 1955; Balogh J.. Lebensgemeinschaften der Landtiere, Bdpst - B., 1958; Kormondy Е. J., Readings in ecology, L., 1965. М. С. Гиляров. Биоценология Биоценоло'гия(от биоценоз и ...логия ), наука, изучающая растительные и животные сообщества в их совокупности, т. е. биоценозы, их строение, развитие, распределение в пространстве и во времени, происхождение. Изучение сообществ организмов в их взаимодействии с неживой природой - предмет биогеоценологии . Биоценометр Биоцено'метр(от биоценоз и ...метр ), прибор для количественного учёта наземных насекомых и других беспозвоночных, применяемый при экологических исследованиях. Б. представляет собой цилиндр или куб без дна, стенки и верх которого затянуты мелкой сеткой или марлей. Применение Б.: пробная площадка размером 0,25-1 м 2быстро (чтобы не ускользнули животные) накрывается Б. Пойманных животных извлекают из Б., умерщвляют, подсчитывают по группам и видам, взвешивают и определяют относительную численность их и массу на единицу площади данного биотопа . Биоциклы Биоци'клы,или жизненные области, три самых крупных подразделения биосферы : суша, море и внутренние водоёмы. Каждый Б. подразделяется на биохоры , включающие значительное число биотопов . Например, биотопы песчаных, глинистых и каменистых пустынь объединяются в биохор пустынь, который вместе с биохорами лесов, степей и др. составляет Б. суши. Термин биологические циклы употребляется в экологии в ином значении. Лит.:Гептнер В. Г., Общая зоогеография, М. -Л., 1936; Наумов Н. П., Экология животных, М., 1955; Вернадский В. И., Химическое строение биосферы Земли и её окружения, М., 1965; Ecological animal geography, N. Y., 1951. И. А. Шилов. Биоэлектрические потенциалы Биоэлектри'ческие потенциа'лы,электрические потенциалы, возникающие в тканях и отдельных клетках человека, животных и растений, важнейшие компоненты процессов возбуждения и торможения.Исследование Б. п. имеет большое значение для понимания физико-химических и физиологических процессов в живых системах и применяется в клинике с диагностической целью ( электрокардиография , электроэнцефалография , электромиография и др.). Первые данные о существовании Б. п. («животного электричества») были получены в 3-й четверти 18 в. при изучении природы «удара», наносимого некоторыми рыбами с электрическими органами при защите или нападении. К этому же времени относится начало исследований итальянского физиолога и врача Л. Гальвани, заложивших основу учения о Б. п. Многолетний научный спор (1791-97) между Л. Гальвани и физиком А. Вольта о природе «животного электричества» завершился двумя крупными открытиями: были получены факты о существовании биоэлектрических явлений в живых тканях и открыт новый принцип получения электрического тока с помощью разнородных металлов - создан гальванический элемент ( вольтов столб ). Правильная оценка наблюдений Гальвани стала возможной лишь после применения достаточно чувствительных электроизмерительных приборов - гальванометров . Первые такие исследования были проведены итальянским физиком К. Маттеуччи (1837). Систематическое изучение Б. п. было начато немецким физиологом Э .Дюбуа-Реймоном (1848), который доказал существование Б. п. в нервах и мышцах в покое и при возбуждении. Но ему не удалось (в силу большой инерционности гальванометра) зарегистрировать быстрые, длящиеся тысячные доли секколебания Б. п. при проведении импульсов вдоль нервов и мышц. В 1886 немецкий физиолог Ю. Бернштейн проанализировал форму потенциала действия; французский учёный Э. Ж. Марей (1875) применил для записи колебаний потенциалов бьющегося сердца капиллярный электрометр; русский физиолог Н. Е. Введенский использовал (1883) для прослушивания ритмических разрядов импульсов в нерве и мышце телефон, а голландский физиолог В. Эйнтховен (1903) ввёл в эксперимент и клиническую практику струнный гальванометр - высокочувствительный и малоинерционный прибор для регистрации электрических токов в тканях. Значительный вклад в изучение Б. п. внесли русские физиологи: В. В. Правдич-Неминский (1913-21) впервые зарегистрировал электроэнцефалограмму, А. Ф. Самойлов (1929) исследовал природу нервно-мышечной передачи возбуждения, а Д. С. Воронцов (1932) открыл следовые колебания Б. п., сопровождающие потенциал действия в нервных волокнах. Дальнейший прогресс в изучении Б. п. был тесно связан с успехами электроники, позволившими применить в физиологическом эксперименте электронные усилители и осциллографы (работы американских физиологов Г. Бишопа, Дж. Эрлангера и Г. Гассера в 30-40-х гг. 20в.). Изучение Б.п. в отдельных клетках и волокнах стало возможным с разработкой микроэлектродной техники . Важное значение для выяснения механизмов генерации Б. п. имело использование гигантских нервных волокон головоногих моллюсков, главным образом кальмара. Диаметр этих волокон в 50 - 100 раз больше, чем у позвоночных животных, он достигает 0,5-1 мм, что позволяет вводить внутрь волокна микроэлектроды, инъецировать в протоплазму различные вещества и т.п. Изучение ионной проницаемости мембраны гигантских нервных волокон позволило английским физиологам А. Ходжкину, А. Хаксли и Б. Катцу (1947-52) сформулировать современную мембранную теорию возбуждения. Различают следующие основные виды Б. п. нервных и мышечных клеток: потенциал покоя, потенциал действия, возбуждающие и тормозные постсинаптические потенциалы, генераторные потенциалы. Потенциал покоя(ПП, мембранный потенциал покоя). У живых клеток в покое между внутренним содержимым клетки и наружным раствором существует разность потенциалов (ПП) порядка 60- 90мв, которая локализована на поверхностной мембране. Внутренняя сторона мембраны заряжена электроотрицательно по отношению к наружной ( рис. 1 ). ПП обусловлен избирательной проницаемостью покоящейся мембраны для ионов К +(Ю. Бернштейн, 1902, 1912; А. Ходжкин и Б. Катц, 1947). Концентрация К +в протоплазме примерно в 50 раз выше, чем во внеклеточной жидкости, поэтому, диффундируя из клетки, ионы выносят на наружную сторону мембраны положительные заряды, при этом внутренняя сторона мембраны, практически не проницаемой для крупных органических анионов, приобретает отрицательный потенциал. Поскольку проницаемость мембраны в покое для Na +примерно в 100 раз ниже, чем для К +, диффузия натрия из внеклеточной жидкости (где он является основным катионом) в протоплазму мала и лишь незначительно снижает ПП, обусловленный ионами К +. В скелетных мышечных волокнах в возникновении потенциала покоя важную роль играют также ионы Cl -, диффундирующие внутрь клетки. Следствием ПП является ток покоя, регистрируемый между поврежденным и интактным участками нерва или мышцы при приложении отводящих электродов. Мембраны нервных и мышечных клеток (волокон) способны изменять ионную проницаемость в ответ на сдвиги мембранного потенциала. При увеличении ПП (гиперполяризация мембраны) проницаемость поверхностных клеточных мембран для Na +и К +падает, а при уменьшении ПП (деполяризация) она возрастает, причём скорость изменений проницаемости для Na +значительно превышает скорость увеличения проницаемости мембраны для К +. Потенциал действия(ПД). Все раздражители, действующие на клетку, вызывают в первую очередь снижение ПП; когда оно достигает критического значения (порога), возникает активный распространяющийся ответ - ПД ( рис. 2 ). Во время восходящей фазы ПД кратковременно извращается потенциал на мембране: её внутренняя сторона, заряженная в покое электроотрицательно, приобретает в это время положительный потенциал. Достигнув вершины, ПД начинает падать (нисходящая фаза ПД), и потенциал на мембране возвращается к уровню, близкому к исходному, - ПП. Полное восстановление ПП происходит только после окончания следовых колебаний потенциала - следовой деполяризации или гиперполяризации, длительность которых обычно значительно превосходит продолжительность пика ПД. Согласно мембранной теории, деполяризация мембраны, вызванная действием раздражителя, приводит к усилению потока Na +внутрь клетки, что уменьшает отрицательный потенциал внутренней стороны мембраны - усиливает её деполяризацию. Это, в свою очередь, вызывает дальнейшее повышение проницаемости для Na +и новое усиление деполяризации и т.д. В результате такого взрывного кругового процесса, т. н. регенеративной деполяризации, происходит извращение мембранного потенциала, характерное для ПД. Повышение проницаемости для Na +очень кратковременно и сменяется её падением ( рис. 3 ), а следовательно, уменьшением потока Na +внутрь клетки. Проницаемость для К +, в отличие от проницаемости для Na +, продолжает увеличиваться, что приводит к усилению потока К +из клетки. В результате этих изменений ПД начинает падать, что ведёт к восстановлению ПП. Таков механизм генерации ПД в большинстве возбудимых тканей. Существуют, однако, клетки (мышечные волокна ракообразных, нервные клетки у ряда брюхоногих моллюсков, некоторые растительные клетки), у которых восходящая фаза ПД обусловлена повышением проницаемости мембраны не для ионов Na +, а для ионов Ca +. Своеобразен также механизм генерации ПД в мышечных волокнах сердца, для которых характерно длительное плато на нисходящей фазе ПД ( рис. 2 , б). Неравенство концентраций ионов К +и Na +(или Ca +) внутри и снаружи клетки (волокна) поддерживается специальным механизмом (т. н. «натриевым насосом» ), выталкивающим ионы Na +из клетки и нагнетающим ионы К +в протоплазму, требующим затраты энергии, которая черпается клеткой в процессах обмена веществ. Амплитуда ПД большинства нервных и мышечных волокон примерно одинакова: 110-120 мв.Длительность ПД варьирует в широких пределах: у теплокровных животных длительность ПД нервных волокон, наиболее быстро проводящих возбуждение, - 0,3-0,4 мсек, у волокон же мышц сердца - 50-600 мсек.В растительных клетках пресноводной водоросли хара ПД продолжается около 20 сек.Характерной особенностью ПД, отличающей его от других форм ответа клетки на раздражение, является то, что он подчиняется правилу «всё или ничего», т. е. возникает только при достижении раздражителем некоторого порогового значения, и дальнейшее увеличение интенсивности раздражителя уже не сказывается ни на амплитуде, ни на продолжительности ПД. Потенциал действия - один из важнейших компонентов процесса возбуждения. В нервных волокнах он обеспечивает проведение возбуждения от чувствительных окончаний ( рецепторов ) к телу нервной клетки и от неё - к синаптическим окончаниям (см. Синапсы ), расположенным на различных нервных, мышечных или железистых клетках. Поступая в эффекторные окончания, ПД вызывает выделение (секрецию) определённой порции специфических химических веществ, т. н. медиаторов , оказывающих возбуждающее или тормозящее влияние на соответствующие клетки. В мышечных волокнах распространяющийся ПД вызывает цепь физико-химических реакций, лежащих в основе процесса сокращения мышц. Проведение ПД вдоль нервных и мышечных волокон осуществляется т. н. локальными токами, или токами действия, возникающими между возбуждённым (деполяризованным) и соседними с ним покоящимися участками мембраны (см. Возбуждение ). Токи действия регистрируются обычными внеклеточными электродами; при этом кривая имеет двухфазный характер: первая фаза соответствует приходу ПД под ближний электрод, вторая - под дальний электрод ( рис. 4 ). Постсинаптические потенциалы(ПСП) возникают в участках мембраны нервных или мышечных клеток, непосредственно граничащих с синаптическими окончаниями. Они имеют амплитуду порядка нескольких мви длительность 10-15 мсек.ПСП подразделяются на возбуждающие (ВПСП) и тормозные (ТПСП). ВПСП представляют собой местную деполяризацию постсинаптической мембраны, обусловленную действием соответствующего медиатора (например, ацетилхолина в нервно-мышечном соединении). При достижении ВПСП некоторого порогового (критического) значения в клетке возникает распространяющийся ПД ( рис. 5 , а, б). ТПСП выражается местной гиперполяризацией мембраны, обусловленной действием тормозного медиатора ( рис. 5 , в). В отличие от ПД, амплитуда ПСП постепенно увеличивается с увеличением количества выделившегося из нервного окончания медиатора. ВПСП и ТПСП суммируются друг с другом при одновременном или последовательном поступлении нервных импульсов к окончаниям, расположенным на мембране одной и той же клетки. Генераторные потенциалывозникают в мембране чувствительных нервных окончаний - рецепторов. Они внешне сходны с ВПСП - их амплитуда порядка нескольких мви зависит от силы приложенного к рецептору раздражения ( рис. 6 ). Когда генераторный потенциал достигает порогового (критического) значения, в соседнем участке мембраны нервного волокна возникает распространяющийся ПД. Ионный механизм генераторных потенциалов ещё недостаточно изучен. Наряду с перечисленными относительно быстро развивающимися Б. п., в нервных клетках, волокнах гладких мышц и некоторых растительных клетках регистрируются также очень медленные колебания мембранного потенциала неизвестной природы, причём на гребне волны деполяризации мембраны часто возникают разряды импульсов. Все Б. п. могут быть зарегистрированы и точно измерены только с помощью внутриклеточных микроэлектродов, позволяющих отводить разности потенциалов между внутренней и наружной сторонами мембраны клетки. При отведении колебаний Б. п. от целых нервов, мышц или мозга с помощью поверхностных электродов регистрируется лишь суммарно потенциал множества синхронно или, чаще, асинхронно работающих клеток. Так, электромиограмма представляет собой результат сложения (интерференции) ПД множества скелетных мышечных волокон; электрокардиограмма - результирующая колебаний электрических потенциалов мышечных волокон различных отделов сердца; электроэнцефалограмма - результат суммации главным образом ВПСП и ТПСП множества клеток различных слоев коры больших полушарий. Регистрация таких интерференционных электрограмм, хотя и не позволяет анализировать колебания Б. п. отдельных клеток, имеет важное значение для суждения о состоянии исследуемого органа в целом. В клинической практике электромиограмму, электрокардиограмму и электроэнцефалограмму регистрируют с помощью электродов, расположенных на коже соответствующих частей тела. Оценка данных, полученных этими методами, основана на сопоставлении изменений характера соответствующей кривой с результатами клинических, физиологических и патологоанатомических исследований. Лит.;Физиология человека, М., 1966; Гальвани Л. и Вольта А., Избр. работы о животном электричестве, М. - Л., 1937; Ходжкин А., Нервный импульс, пер. с англ., М., 1965; Экклс Дж., Физиология нервных клеток, пер. с англ., М., 1959; его же, Физиология синапсов, М., 1966; Катц Б., Нерв, мышца и синапс, пер. с англ., М., 1968; Ходоров Б. И., Проблема возбудимости, Л., 1969. Б. И. Ходоров. 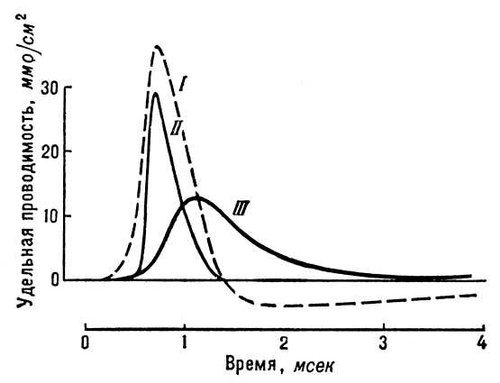
Рис. 3. Изменения натриевой и калиевой проводимости мембраны нервного волокна во время генерации потенциала действия (I). Изменения проводимости пропорциональны изменениям проницаемости для Na +(II) и К +(III). 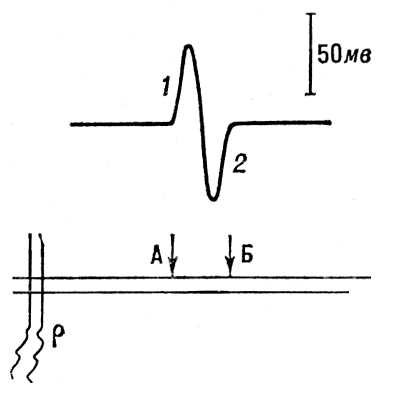
Рис. 4. Регистрация распространения потенциала действия вдоль нервного волокна. А, Б - внеклеточные электроды; р - раздражающие электроды. Вверху - отклонение луча осциллографа под влиянием волны возбуждения; 1 - волна возбуждения под электродом А; 2 - под электродом Б. 
Рис. 1. Схема измерений мембранного потенциала покоя с помощью внутриклеточного стеклянного микроэлектрода (М). Второй электрод (И) помещен в омывающую клетку жидкость. 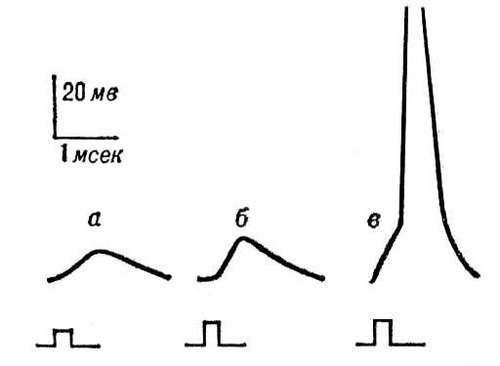
Рис. 6. Генераторные потенциалы: увеличение амплитуды при усилении раздражения рецептора (а - в). При достижении порогового значения (в) генераторный потенциал вызвал в чувствительном нервном волокне потенциал действия. 
Рис. 5. Постсинаптические потенциалы: а - подпороговые ВПСП, возникающие в нервной клетке в ответ на раздражения соответствующих нервных волокон; б - ВПСП, достигший порогового значения, достаточного для возникновения ПД; в - ТПСП, вызванный раздражением тормозных нервных волокон. 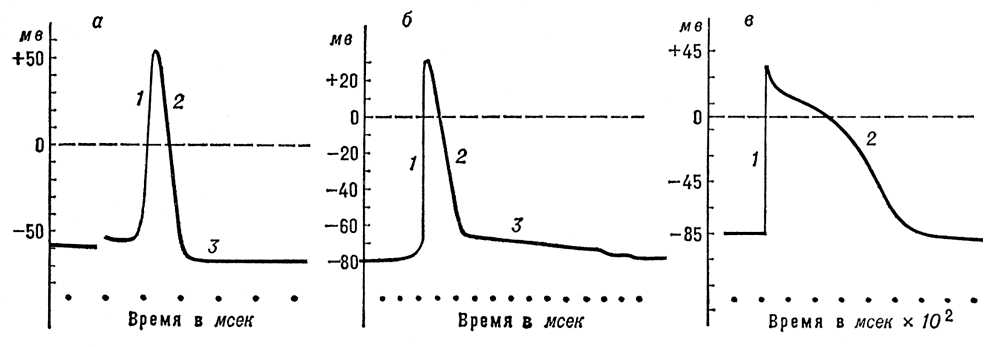
Рис. 2. Потенциалы действия, зарегистрированные с помощью внутриклеточных микроэлектродов: а - гигантского аксона кальмара; б - скелетного мышечного волокна; в - волокна мышцы сердца собаки; 1 - восходящая фаза ПД; 2 - нисходящая фаза; 3 - следовая гиперполяризация (а) и следовая деполяризация (б). Биоэлектрические явления Биоэлектри'ческие явле'ния,см. Биоэлектрические потенциалы . Биоэнергетика Биоэнерге'тика,биологическая энергетика, изучает механизмы преобразования энергии в процессах жизнедеятельности организмов. Иначе говоря, Б. рассматривает явления жизнедеятельности в их энергетическом аспекте. Методы и подходы к изучаемым явлениям, применяемые в Б., - физико-химические, объекты и задачи - биологические. Т. о., Б. стоит на стыке этих наук и является частью молекулярной биологии , биофизики и биохимии . Началом Б. можно считать работы немецкого врача Ю. Р. Майера , открывшего закон сохранения и превращения энергии (1841) на основе исследования энергетических процессов в организме человека. Суммарное изучение процессов, являющихся источниками энергии для живых организмов (см. Дыхание , Брожение ), и энергетического баланса организма, его изменений при различных условиях (покой, труд разной интенсивности, окружающая температура) долгое время являлось основным содержанием Б. (см. Основной обмен , Теплоотдача , Теплопродукция ). В середине 20 в., в связи с общим направлением развития биологических наук, центральное место в Б. заняли исследования механизма преобразования энергии в живых организмах. Все исследования в области Б. основываются на единственно научной точке зрения, согласно которой к явлениям жизни полностью применимы законы физики и химии, а к превращениям энергии в организме - основные начала термодинамики . Однако сложность и специфичность биологических структур и реализующихся в них процессов обусловливают ряд глубоких различий между Б. и энергетикой неорганического мира, в частности технической энергетикой. Первая фундаментальная особенность Б. заключается в том, что организмы - открытые системы , функционирующие лишь в условиях постоянного обмена веществом и энергией с окружающей средой. Термодинамика таких систем существенно отличается от классической. Основополагающее для классической термодинамики понятие о равновесных состояниях заменяется представлением о стационарных состояниях; второе начало термодинамики (принцип возрастания энтропии ) получает иную формулировку в виде Пригожина теоремы . 1, 2, 3, 4, 5, 6, 7, 8, 9, 10, 11, 12, 13, 14, 15, 16, 17, 18, 19, 20, 21, 22, 23, 24, 25, 26, 27, 28, 29, 30, 31, 32, 33, 34, 35, 36, 37 |
|||||||