Физиология размножения и репродуктивная патология собак
ModernLib.Net / Биология / Дюльгер Георгий / Физиология размножения и репродуктивная патология собак - Чтение
(стр. 2)
|
Автор:
|
Дюльгер Георгий |
|
Жанр:
|
Биология |
|
-
Читать книгу полностью
(317 Кб)
- Скачать в формате fb2
(536 Кб)
- Скачать в формате doc
(112 Кб)
- Скачать в формате txt
(108 Кб)
- Скачать в формате html
(542 Кб)
- Страницы:
1, 2, 3, 4, 5, 6, 7, 8, 9, 10, 11
|
|
Гормоны яичников не оказывают решающего влияния на половую дифференцировку репродуктивной системы самок в период преднатального развития. При врожденном отсутствии гонад или в результате их удаления на стадии индифферентной гонады развиваются половые органы только женского типа. При гермафродитизме (аномалии полового развития, когда гонады дифференцируются сразу в двух направлениях — овариальном и тестикулярном) развитие внутренних и наружных гениталий зависит от наличия и активности тестостерон-секретирующих клеток в гонаде смешанного типа. В постнатальном периоде во время полового созревания формируется суточная цикличность и увеличивается выделение гонадотропинов, под влиянием которых возрастает секреция эстрогенных гормонов яичников, устанавливается взаимодействие между яичниками и гипоталамо-гипофизарной системой. Фолликулогенез носит незавершенный характер. Фолликулы дегенерируют на разных стадиях своего развития. Процесс дегенерации третичных фолликулов называется атрезией. Генеративную активность яичники начинают проявлять при наступлении половой зрелости. Созревание половых клеток носит циклический характер: в течение каждого полового цикла несколько третичных фолликулов вступают в финальную стадию своего развития, созревают и овулируют. Яйцеклетка овулирует на стадии ооцита первого порядка (не содержит полярных телец). Она правильной шарообразной формы, ее диаметр 1,2?10
-5мм, а вместе с прозрачной оболочкой 1,56?10
-5мм (Hoist P.A. et al., 1971). Ооцит созревает в верхней трети яйцепроводов, претерпевая два мейотических деления. Первое мейотическое деление инициируется преовуляторным выбросом лютеинизирующего гормона (ЛГ) и приводит к образованию ооцита второго порядка и отделению в перивителлиновое пространство первого полярного тельца, содержащего незначительное количество цитоплазмы и лишние хромосомы. При проникновении в яйцеклетку спермия ооцит второго порядка претерпевает второе деление мейоза, в результате которого образуются одна зрелая яйцеклетка с гаплоидным набором хромосом, способная к оплодотворению, и второе полярное тельце.
1.4. НЕЙРОЭНДОКРИННАЯ РЕГУЛЯЦИЯ ПОЛОВЫХ ПРОЦЕССОВ
Важнейшая функция нервной системы — управление деятельностью целостного организма при помощи нервных и гуморальных сигналов на основании сбора, анализа и интеграции информации, исходящей от отдельных частей организма и из окружающей среды. По топографическому признаку нервную систему подразделяют на центральную и периферическую.
Центральная нервная система (ЦНС)
включает в себя спинной и головной мозг,
периферическая
— спинномозговые и черепные нервы, их ветви и сплетения.
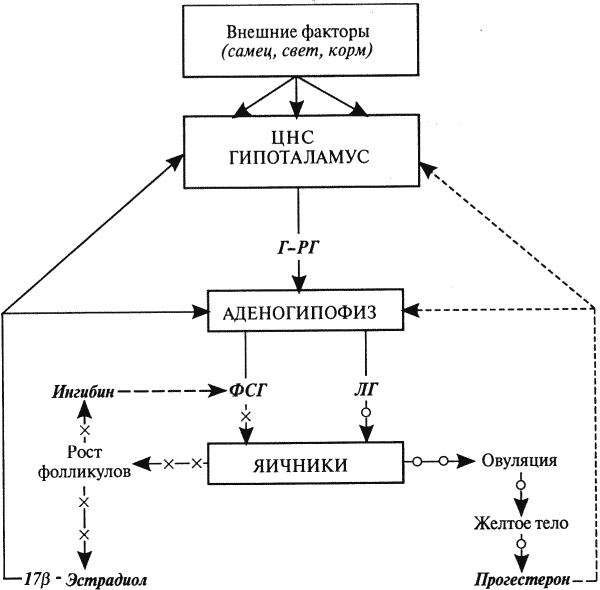
Рис. 8. Схема нейроэндокринной регуляции половых процессов у самок
[Сплошными линиями показана прямая и положительная обратная связь (стимуляция), прерывистыми — отрицательная (блокирование)]: гормон гипоталамуса Г-РГ стимулирует выделение из аденогипофиза ФСГ и ЛГ; ФСГ активирует рост и развитие фолликулов и выработку ими эстрогенов и ингибина — ингибин избирательно блокирует секрецию ФСГ; эстрогены, воздействуя на органы-мишени (ЦНС, вторичные половые органы), индуцируют проявление течки, полового возбуждения и охоты; в конце проэструса и в начале эструса, когда концентрация прогестерона низкая, преовуляторный пик Пр-эстрадиола инициирует циклический выброс Г-РГ, ФСГ и ЛГ; преовуляторный пик ЛГ индуцирует созревание преовуляторных фолликулов, их овуляцию, базальная секреция ЛГ — формирование желтых тел и выработку ими прогестерона; прогестерон через механизмы отрицательной обратной связи контролирует секрецию Г-РГ, ФСГ и ЛГ: высокие его концентрации блокируют, а низкие стимулируют выделение этих гормонов
По функциональному признаку различают соматическую и вегетативную нервную систему.
Соматическая система
иннервирует органы тела (сому) и связывает организм с внешней средой при помощи органов чувств, кожной чувствительности и движения. Центры соматической системы находятся в ЦНС, высший отдел которой — кора головного мозга — контролирует высшую нервную деятельность.
Вегетативная, или автономная, система
, обеспечивающая иннервацию органов и систем организма, в состав которых входят гладкие мышечные волокна и железистый эпителий (органы пищеварения, дыхания, кровоснабжения, выделения, размножения и внутренней секреции), включает в себя симпатическую и парасимпатическую части. Центры симпатической нервной системы расположены в грудопоясничном отделе спинного мозга, парасимпатической — в стволе головного мозга и крестцовом отделе спинного мозга. Нервный импульс в периферических синапсах симпатической системы передается при помощи норадреналина, парасимпатической — ацетилхолина. Симпатическая и парасимпатическая части координируют работу внутренних органов, оказывая на них противоположное действие, например, у самок возбуждение b-адренорецептеров симпатической системы способствует расслаблению матки, их блокада или же возбуждение холинорецептеров парасимпатической системы, наоборот, стимулирует сокращение органа. У самцов симпатическая часть стимулирует рефлекс эякуляции, парасимпатическая — эрекции. Проявление полового цикла и половых рефлексов зависит от взаимодействия нервной и эндокринной систем. Схема нейроэндокринной регуляции половых процессов у самок показана на рисунке 8, половых процессов у самцов — на рисунке 9.
Гипоталамус
— надбугорная область промежуточного мозга, является одновременно нервным образованием и эндокринной железой. Вырабатывает окситоцин, вазопрессин и еще 10 гипофизотропных нейрогормонов, семь из которых оказывают на переднюю долю гипофиза стимулирующее действие (либерины), три — ингибирующее (статины). В регуляции воспроизводительной функции принимают участие окситоцин, пролактостатин, гонадолиберин и кортиколиберин.
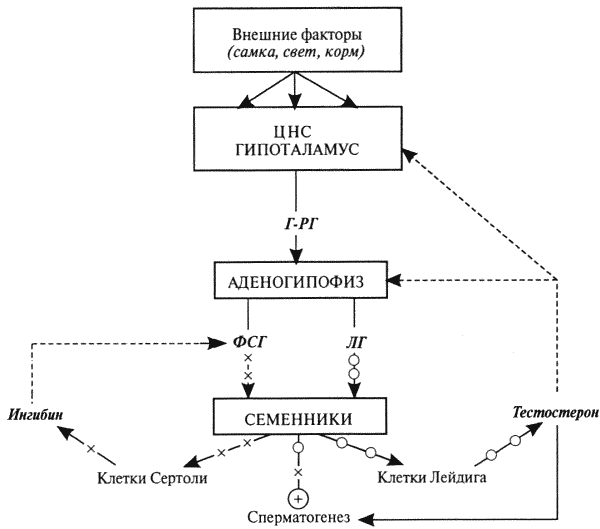
Рис. 9. Схема нейроэндокринной регуляции половых процессов у самцов
[Сплошными линиями показана прямая связь (стимуляция), прерывистыми — отрицательная обратная связь (блокирование)]: гормон гипоталамуса Г-РГ стимулирует выделение из аденогипофиза ФСГ и ЛГ; ЛГ стимулирует выработку клетками Лейдига гормона тестостерона; тестостерон поддерживает сперматогенез, половое влечение и через механизмы отрицательной обратной связи контролирует секрецию Г-РГ, ФСГ и ЛГ: высокие его концентрации блокируют, а низкие — стимулируют выделение этих гормонов; ФСГ стимулирует выработку клетками Сертоли гормона ингибина; инги-бин через механизмы обратной связи подавляет секрецию ФСГ
Окситоцин
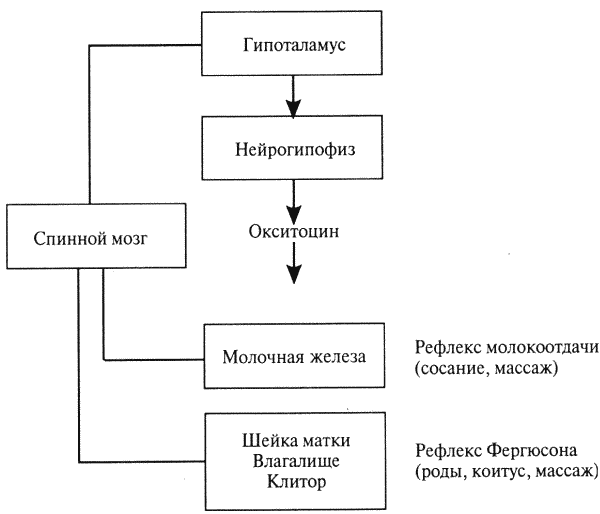
представляет собой нанопептид; вырабатывается гипоталамусом и аккумулируется в задней доле гипофиза (нейро-гипофизе). Окситоцин стимулирует сократительную активность матки, принимает участие в родовом акте и способствует отдаче секрета молочных желез во время акта сосания. У самцов окситоцин, по-видимому, вызывает перистальтические сокращения в спермиопроводах. Секреция гормона в организме регулируется нейрорефлекторным путем (рис. 10).
Пролактостатин
, или пролактинингибирующий фактор (ПИФ), блокирует секрецию пролактина передней долей гипофиза (аденогипофизом). Секрецию пролактостатина стимулирует дофамин. На применении агонистов дофамина основано подавление лактации в ветеринарной и медицинской практике.
Гонадолиберин
, или люлиберин, гонадотропин-рилизинг-фактор, гонадотропин-рилизинг-гормон Г-РГ, ЛГ-РГ, ФСГ/ЛГ-РГ, — декапептид, регулирующий синтез и секрецию гонадотропных гормонов гипофиза —
фоллитропина
(фолликулостимулирующий гормон, или ФСГ) и
лютропина
(лютеинизирующий гормон, или ЛГ). ФСГ и ЛГ по химическому составу представляют собой глю-копротеиды. ФСГ стимулирует у самок рост и развитие фолликулов, у самцов — сперматогенез; ЛГ — созревание преовуляторных фолликулов, их овуляцию, образование желтого тела и выработку прогестерона. Вместе с ФСГ лютропин инициирует также секрецию эстрогенов третичными фолликулами. У самцов ЛГ стимулирует продукцию клетками Лейдига мужского полового гормона тестостерона.
Рис. 10. Нейрорефлекторные пути высвобождения окситоцина гипоталамо-гипофизарной системой
Пролактин
(ПРЛ), лактогенный или лютеотропный гормон — полипептид, вырабатываемый аденогипофизом; у самок стимулирует процесс образования молока, поддерживает лактацию и во второй половине беременности проявляет лютеотропные свойства. У самцов его действие неизвестно. Характерная особенность ПРЛ — органы-мишени пролактина (молочная железа, желтое тело) не синтезируют гормоны, тормозящие его секрецию (отсутствие обратной связи). Существует два типа секреции гонадотропинов: тоническая и циклическая. Тоническая секреция непрерывна, ее регистрируют у самцов и самок на протяжении всей жизни. Циклическая секреция ФСГ и ЛГ предшествует овуляции, ее регистрируют у половозрелых самок. Преовуляторный выброс ЛГ у спонтанно овули-рующих животных инициируется преовуляторным пиком эстрадиола, у рефлекторно овулирующих (кошки, кролики, верблюды) — половым актом. Эпифиз, или шишковидная железа, — верхний мозговой придаток, относящийся к структурам промежуточного мозга. Вырабатывает нейросекрет
мелатонин
, характеризующийся (в зависимости от вида животного) анти- и прогонадальной активностью: способностью тормозить или, наоборот, стимулировать секрецию гонадолиберина. Отличительная особенность эпифиза — зависимость его секреторной активности от освещенности (фотопериодизм). Активность железы возрастает в темное время суток. Нейрогормон эпифиза контролирует суточный гормональный ритм в организме. У многих моно- (волки, койоты, шакалы, дикие собаки Динго и др.) и полициклических животных (представители семейства кошачьих, овцы, лошади и др.) световой фактор служит основным климатическим регулятором сезонности размножения. Роль мелатонина у собак остается невыясненной, так как собаки проявляют половую активность вне зависимости от сезона года. Физиологическая роль половых гормонов в организме самок и самцов исключительно многообразна. Эндокринные части яичников продуцируют эстрогены, прогестерон и ингибин, семенников — тестостерон и ингибин.
Эстрогены
— половые гормоны стероидной природы, состоят из 18 атомов углерода (С
18). Вырабатываются растущими и созревающими третичными фолликулами и плацентой. Различают три фракции эстрогенов — эстрадиол, эстрон и эстриол. Эстрадиол, наиболее активный из перечисленных, представляет собой первичный эстроген, который может превращаться в эстрон и эстриол. В организме самок эстрогены ответственны за развитие вторичных половых признаков и выводных протоков молочных желез. Эстрогены индуцируют течку, половое возбуждение и охоту, вызывают пролиферацию эндометрия, миометрия, эпителия слизистой оболочки влагалища и его преддверия, усиливают кровоснабжение половых органов, способствуют раскрытию цервикального канала, стимулируют сократительную активность матки, маточных труб и принимают активное участие в родовой деятельности, через механизмы положительной обратной связи индуцируют Преовуляторный выброс ЛГ у спонтанно овулирующих животных.
Прогестерон
— стероидный гормон (C
21). Вырабатывается желтым телом полового цикла и беременности, а также плацентой. У собак желтое тело — основной продуцент прогестерона на протяжении всего периода беременности. Овариоэктомия приводит к прерыванию беременности на любой ее стадии. Прогестерон переводит эндометрий в секреторно активное состояние, подготавливает его к прикреплению зародыша, поддерживает в матке условия, необходимые для развития эмбриона и плода, блокирует сократительную активность матки, вызывает закрытие цервикально-го канала, подавляет созревание фолликулов, проявление течки, полового возбуждения и охоты, стимулирует развитие альвеол молочной железы и тормозит секрецию ЛГ. 2. Участие гормонов в регулировании репродуктивной функции у собак
Ингибин
— пептидный гормон, вырабатываемый фолликулярным эпителием третичных фолликулов и клетками Сертоли. Тормозит секрецию ФСГ в организме самок и самцов.
Релаксин
— пептидный гормон, вырабатываемый желтым телом в конце беременности. Подготавливает материнский организм к родовому акту, вызывая расслабление связок таза, мышц шейки матки, влагалища и его преддверия.
Тестостерон
— мужской половой гормон стероидной природы (С
19), вырабатываемый в семенниках, яичниках, коре надпочечников. В семенниках тестостерон синтезируется в клетках Лейдига, в яичниках — в третичных фолликулах, являясь лишь промежуточным этапом биосинтеза. Физиологическая роль тестостерона в эмбриональный период — участие в половой дифференцировке организма. Тестостерон необходим для развития вторичных половых признаков, поддержания сперматогенеза, стимуляции полового влечения и регуляции секреции ЛГ у самцов. В регуляции половых процессов принимают также участие материнские и фетальные глюкокортикостероиды и простагландин Фзц.
Глюкокортикоиды
— стероидные гормоны, вырабатываемые надпочечниками матери и плода. Кортизол (гормон стресса) — наиболее активный и принципиальный глюкокортикоидный гормон, принимающий участие в регуляции родового процесса и лактогенеза. В инициации родового акта фетальному кортизолу, по-видимому, принадлежит ведущая роль. Секрецию глюкокортикоидов контролируют
кортиколиберин
и
адренокортикотропный гормон
, или АКТГ. Кортиколиберин — пептидный гормон гипоталамуса, стимулирует секрецию полипептидного гормона аденогипофиза АКТГ, характеризующегося кортикотропной активностью.
ПростагландинФ
2
a(ПгФ
2
a) — биологически активное соединение с гормональными свойствами, производное полиненасыщенных жирных кислот. Вырабатывается многими клетками и тканями и принимает участие в разнообразных процессах и реакциях организма. У многих домашних животных ПгФ
2
aслужит маточным лютеолитическим фактором (вызывает регрессию желтого тела полового цикла и беременности) и принимает активное участие в инициации и поддержании родового процесса. Желтые тела полового цикла и беременности у собак и кошек мало чувствительны к нему. Аборт у животных данных видов обычно удается вызвать только после многократного применения ПгФ
2
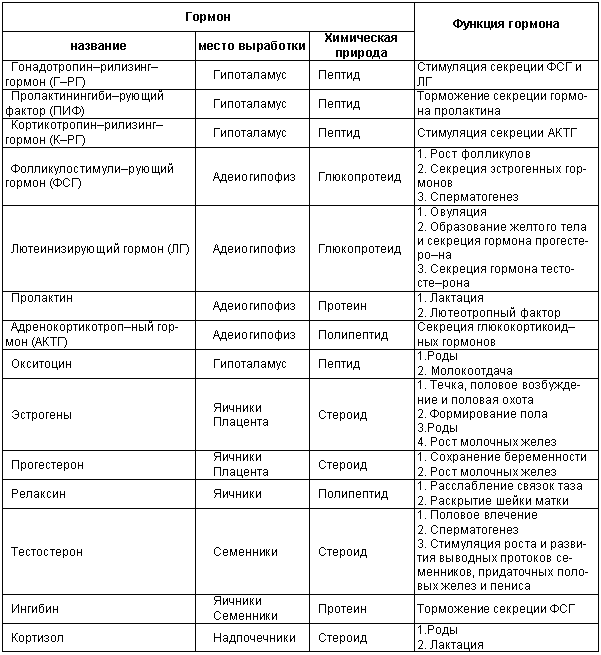
a, начиная с 5-го дня диэструса. На нейроэндокринную регуляцию половой функции собак существенное влияние оказывает общение с противоположным полом (особенно большое значение это имеет для рефлекторно овулирующих животных — кроликов, верблюдов, лам, представителей семейства кошачьих). Координацию полового поведения самок и самцов обеспечивают органы чувств, кожная чувствительность и движение. Важным внешним фактором, влияющим на все функции организма собак, служит корм. Недостаточное, избыточное и (или) неполноценное кормление не только снижает воспроизводительную способность животных, но часто бывает причиной их бесплодия. Важнейшие репродуктивные функции гормонов гипоталамуса, гипофиза, гонад, плаценты и надпочечников указаны в таблице 2.
1.5. ПОЛОВАЯ И ФИЗИОЛОГИЧЕСКАЯ ЗРЕЛОСТЬ
Половая зрелость
— это возраст, начиная с которого самцы и самки способны участвовать в процессе полового размножения: продуцировать соответствующие половые клетки (спермии, ооциты) и спариваться. Половая активность у самок носит циклический характер. Готовность к спариванию они демонстрируют только в строго фиксированный период полового цикла — во время половой охоты. Овуляция у собак — спонтанная и привязана к началу половой охоты. Выделение спермы (спермиев и плазмы спермы) из половых органов самцов и введение ее в половые органы самок происходят во время полового акта.
Сроки наступления зрелости у самок определяют по дате проявления первого полового цикла. Обычно первый половой цикл регистрируют в возрасте 6…12 мес. У самцов половая зрелость наступает примерно на 1…1,5 мес позднее, чем у самок. Собаки средних и карликовых пород становятся половозрелыми раньше, чем собаки крупных пород. На сроки наступления половой зрелости существенное влияние оказывают состояние здоровья животного, условия кормления и содержания, общение с противоположным полом.
Половая зрелость наступает обычно раньше, чем заканчиваются основной рост, структурное и физиологическое развитие животного, обеспечивающие его высокую плодовитость, нормальное функционирование организма в период беременности, родов и лактации, рождение здорового приплода.
Физиологическая зрелость — возраст, начиная с которого самцов и самок целесообразно использовать для получения приплода. Физиологическая зрелость у собак по времени совпадает, как правило, с достижением размеров тела, характерных для взрослого животного, и проявлением у самок 2…3 половых циклов. Самки большинства пород становятся физиологически зрелыми обычно в возрасте 1,5 лет, самцы — 2 лет.
1.6. ПОЛОВОЙ ЦИКЛ
По ритму полового цикла животных общепринято подразделять на моно- (с одним половым циклом в год) и полициклических, у которых несколько циклов повторяются с определенной для каждого вида животного периодичностью на протяжении всего года или определенного сезона года. Собаки занимают как бы промежуточное положение: на протяжении года у них регистрируют от одного до двух и даже трех половых циклов. По этой причине одни авторы относят собак к моно-, другие — к дициклическим животным. По многим признакам собак следует определять как моноциклических животных. Во-первых, дикие представители семейства собачьих (волки, койоты, шакалы, дикие собаки Динго), с которыми домашняя собака имеет много общих черт и легко скрещивается, — это типичные моноциклические животные, единственный половой цикл которых проявляется в строго фиксированный сезон года. Во-вторых, в половом цикле собак есть характерная для моноциклических животных особенность: течку наблюдают после хорошо выраженного анэстрального периода. В процессе одомашнивания половая цикличность собак изменилась: теперь у них регистрируют несколько половых циклов в течение года и вне зависимости от сезона (Sokolowski J.H., 1977). Исключение составляют собаки породы басенджи, единственный половой цикл которых проявляется обычно осенью. Указанная особенность, по-видимому, обусловлена действием простого рецессивного гена, поскольку при скрещивании собак породы басенджи с собаками других пород у одних потомков отмечают один половой цикл в год, у других — несколько (McDonald L.E., 1980). В повседневной практике половую активность самок учитывают по межэстральному периоду, так как определить продолжительность полового цикла и, в частности, время его окончания можно только с помощью лабораторных методов исследований: по динамике прогестерона в крови и картине влагалищных мазков. Продолжительность межэстрального периода сильно варьирует и зависит от породы собаки, но не от ее размеров (табл. 3). 3. Продолжительность межэстрального периода у собак некоторых пород (Sokolowski J.H., 1977)
Беременность не оказывает существенного влияния на продолжительность межэстрального периода: у самок породы бигль его продолжительность при наступлении беременности в среднем составляет 245,1 дня, при ее отсутствии — 235,8 дня (Sokolowski J.H., 1977). Половой цикл собак уникален, он по многим признакам отличается от цикла сельскохозяйственных и лабораторных животных. Наступлению половой охоты у собак предшествуют геморрагические выделения, которые возникают в результате диапедеза (просачивания клеток крови через стенку кровеносного сосуда). Яйцеклетка овулирует в начале половой охоты незрелой (на стадии ооцита первого порядка) и способность к оплодотворению приобретает через 2…3 дня после попадания в яйцевод. Функционально активное желтое тело образуется к моменту окончания половой охоты. У большинства животных других видов желтое тело формируется в конце половой охоты или после ее окончания, и этот период принято выделять в самостоятельную стадию полового цикла (метэструс, или стадию торможения). После регрессии желтого тела полового цикла и беременности наступает анэструс (период полового покоя). Видимо, логично делить межэстральный период собак на следующие четыре стадии: проэструс, эструс, диэструс и анэструс. Стадию метэструса у собак следует рассматривать как постовуля-ционную фазу эструса. Бессмысленно ее выделять в самостоятельную стадию, поскольку период овуляции и образования желтого тела у собак не удается диагностировать клиническими методами и он представляет для исследователей только академический интерес. Благодаря делению межэстрального периода на проэструс, эструс, диэструс и анэструс можно избежать путаницы в научной терминологии и обозначить соответствующие стадии полового цикла у животных разных видов одними и теми же терминами. Обзор терминов и критериев начала и окончания стадий полового цикла и анэструса приведен в таблице 4. 4. Терминология и критерии начала и окончания стадий полового цикла и периода полового покоя у собак по данным разных авторов
Примечание: а — время овуляции; б — время окончания половой охоты; в — время окончания функционирования желтого тела полового цикла; г — время завершения регенерации эндометрия.
Проэструс
— стадия подготовки половых органов и всего организма самки к половому акту, продолжительность которой составляет в среднем 9 (2…16) дней. Проэструс у собак хорошо выражен, его легко диагностировать по признакам течки и полового возбуждения. ФСГ стимулирует развитие преовуляторных фолликулов. Про-гестагенная активность яичников находится на самом низком уровне. Концентрация прогестерона в плазме периферической крови ниже 1?10
-9г/мл. Эстрогенная активность яичников по мере развития преовуляторных фолликулов возрастает. Максимальную концентрацию эстрогенов (преовуляторный пик 17?-эстрадиола) в плазме периферической крови регистрируют в конце проэструса или в начале эструса. Эстрогенные гормоны, воздействуя на ЦНС и половые органы, обусловливают развитие признаков течки и полового возбуждения. Течка — процесс выделения слизи из половых органов самки. У собак течка характеризуется следующими признаками: половая петля (вульва) увеличена в размере, набухшая; слизистые оболочки влагалища и его преддверия гиперемированы и отечны; сократительная активность матки повышена. Все слои стенки матки утолщены и обильно кровоснабжены. Шейка матки расслаблена, цервикальный канал раскрыт. Чрезмерная васкуляризация эндометрия приводит к диапедезу клеток крови в просвет матки и появлению геморрагических выделений, которые к концу проэструса становятся более светлыми и менее обильными. Существенные изменения претерпевает эпителий влагалища и его преддверия. Толщина эпителиального слоя резко возрастает, поверхностные эпителиальные клетки ороговевают. Ороговевшие клетки выполняют защитную функцию. Они очень устойчивы к бактериальной инвазии, действию химических и физических факторов. В мазках, взятых из краниальной части влагалища в период проэструса, обнаруживают вариабельное количество лейкоцитов, много эритроцитов и ороговевших эпителиальных клеток без ядра или с пикнотическим ядром.
Половоевозбуждение— изменение в поведении животного. В период проэструса у самок появляется жажда, извращается чутье, они становятся беспокойными, раздражительными, плохо выполняют команды, могут убежать от владельца; обычно часто и в разных местах мочатся — как бы метят территорию, выделяя с мочой ароматические (пахучие) вещества — половые феромоны, привлекающие самцов. Самцы проявляют повышенный «интерес» к самкам с признаками течки и полового возбуждения и беспрерывно преследуют их. «Интерес» самок к самцам обычно возникает только в конце проэструса: самки становятся игривыми, но не допускают садки самца и полового акта.
Эструс
— период половой охоты: положительной сексуальной реакции самки на самца. Половая охота у собак длится в среднем 9 (2…16) дней. Выделения из половой щели, как правило, светлые или со следами крови. Половые губы становятся менее отечными и более ригидными. Самка проявляет повышенный «интерес» к самцам, обнюхивает их половые органы; затем выбирает самца-фаворита, допускает садку и коитус. В начале эструса у самок происходит циклический выброс ЛГ, инициируемый преовуляторным пиком 17? — эстрадиола. Яйцеклетка обычно овулирует на 2…5-й день половой охоты: примерно через 30 ч после того как концентрация ЛГ достигнет максимального значения. На месте овулировавших фолликулов образуются желтые тела, которые становятся функционально активными к концу эструса. После овуляции прогестагенная активность яичников возрастает, а эстрогенная, наоборот, снижается. Во влагалищных мазках в начале эструса отмечают единичные эритроциты и лейкоциты и резкое преобладание ороговевших эпителиальных клеток над эпителиальными клетками, содержащими интактное ядро. Наиболее высокий индекс ороговения во влагалищных мазках регистрируют в период максимальных концентраций ЛГ (преовуляторный пик ЛГ).
Диэструс
— период функционирования желтого тела полового цикла. Начинается с момента окончания половой охоты и заканчивается регрессией желтого тела. Продолжительность диэструса может сильно варьировать (от 40 до 84 дней) и в среднем составляет 60 дней. После окончания половой охоты у самок достаточно быстро исчезают признаки течки и полового возбуждения. Животные становятся спокойными. Во влагалищных мазках преобладают промежуточные и парабазальные эпителиальные клетки, содержащие ядро. Диэструс — самая активная часть лютеиновой фазы полового цикла, начинающаяся сразу же после овуляции. Концентрация прогестерона в плазме периферической крови увеличивается и на 30-й день после проявления преовуляторного пика ЛГ достигает максимальных значений — 25…30?10
-9г/мл (Concannon P. W., 1986). Затем содержание прогестерона начинает медленно снижаться. Механизм регрессии желтого тела полового цикла у собак не выяснен.
Анэструс
— период полового покоя. Его продолжительность может сильно варьировать и в среднем составляет 125 дней. В этот период яичники мало активны. Рост фолликулов носит абортивный (незавершенный) характер. Третичные фолликулы на разных стадиях своего развития подвергаются атрезии. Идет активный лизис желтых тел предыдущего полового цикла. В эндометрии восстанавливается (регенерирует) эпителиальный слой. Во влагалищных мазках встречаются только парабазальные и промежуточные эпителиальные клетки.
1.7. ПОЛОВОЙ АКТ
Половой акт, или совокупление, коитус, копуляция, — совокупность половых рефлексов, обеспечивающих выведение спермы из полового аппарата самца и введение ее в половые пути самки. В половом акте различают следующие рефлексы: приближения, эрекции, обнимательный, совокупительный и эякуляции.
Рефлекс приближения
— совокупность поведенческих реакций, координирующих сексуальное поведение разнополых особей при спаривании.
Рефлекс эрекции
— сильное наполнение кровью и увеличение полового члена в размере, обеспечивающие его выведение из препуция и возможность введения в половые пути самки. Этот рефлекторно-сосудистый акт проявляется у самок набуханием клитора и гиперемией слизистой оболочки влагалища и его преддверия.
Обнимательный рефлекс
— принятие самкой и самцом позы для спаривания. Одновременно с обнимательным проявляется и совокупительный рефлекс.
Совокупительный рефлекс
— введение и фрикция полового члена самца во влагалище самки, приводящие к раздражению эрогенных зон их гениталий и наступлению эякуляции.
Рефлекс эякуляции
— выведение спермы (эякулята) из полового аппарата самца. Эякуляция, так же как и эрекция, это нейрорефлекторный акт. Все половые рефлексы по своей природе врожденные (безусловные) В процессе их становления и приобретения животными сексуального опыта на безусловные рефлексы наслаиваются условные. Одни из них способствуют развитию полноценных половых рефлексов, другие, наоборот, усиливая или ослабляя их проявление, приводят к возникновению анормального полового поведения. Сексуальное поведение животных координируют органы чувств кожная чувствительность и движение. В период проэструса и эструса самки выделяют половые феромоны и в частности, метил-п-гидроксибензоат, стимулирующие половые рефлексы самцов. В начале проэструса самки ведут себя пассивно или же демонстрируют по отношению к самцам агрессивное поведение, в конце проэструса начинают проявлять к ним сексуальный интерес. Самцы следуют за самкой с признаками течки и полового возбуждения, выстроившись по иерархическому рангу Если самка позволяет, то самец-фаворит начинает активно обнюхивать ее, облизывать наружные половые органы и другие участки ее тела, кладет голову, одну или две передние лапы ей на спину, предпринимает попытки совершить половой акт. При наступлении половой охоты самка демонстрирует готовность к спариванию. Обнюхивает половые органы самца, принимает позу для спаривания: стоит неподвижно, отводит в сторону хвост, подтягивает кверху половую петлю. Самец вспрыгивает на самку, обхватывает передними лапами ее за бока и совершает толкательные движения тазом, обеспечивающие введение полового члена во влагалище (рис. 11). 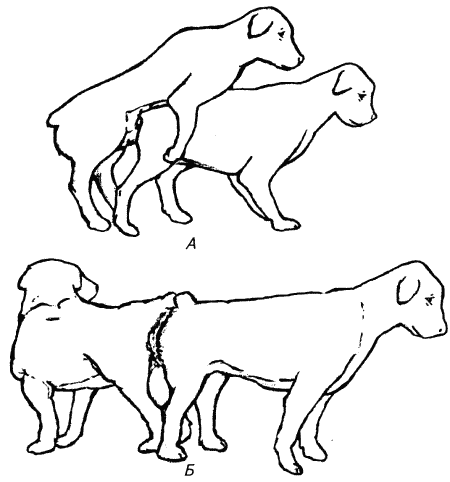 При полном введении пениса рефлекторно сжимается сфинктер преддверия влагалища, что приводит к сильному набуханию луковицы полового члена и сцеплению (склещиванию) половых органов животных. Самец спрыгивает с самки, и животные принимают позу «хвост к хвосту». Сцепление половых органов (половой замок, вязка) продолжается 5…45 мин. Выделение спермы сопровождается ритмичными движениями корня хвоста и волнообразными сокращениями стенки мочеполового канала. Сперма выделяется во влагалище в виде трех фракций: первая фракция выполняет функцию смазки, вторая содержит спермии, третья обеспечивает проталкивание спермиев в полость матки.
Страницы:
1, 2, 3, 4, 5, 6, 7, 8, 9, 10, 11
|
|