 |
|
Популярные авторы:: Горький Максим :: Чехов Антон Павлович :: Раззаков Федор :: БСЭ :: Толстой Лев Николаевич :: Азимов Айзек :: Лондон Джек :: Грин Александр :: Борхес Хорхе Луис :: Лем Станислав Популярные книги:: Справочник по реестру Windows XP :: На страже :: Брат мой... :: Он убил мою собаку :: Приключения Иля :: Загадка для «Секретной семерки» (Тайна пропавшей скрипки, Тайна огородного пугала) :: Чужое :: Джубал Сэкетт :: Патруль :: Английский подснежник |
Большая Советская Энциклопедия (ПО)ModernLib.Net / Энциклопедии / БСЭ / Большая Советская Энциклопедия (ПО) - Чтение (стр. 33)
генов - реализаторов П. Именно эти гены открывают путь в одном случае генам, способствующим образованию женского П., в другом - генам, обусловливающим развитие мужского П. При генетическом определении П. по типу X0, XX реализаторы женского П. локализованы в Х-хромосомах, а мужского - в аутосомах. При сочетании одной дозы реализаторов женского П., локализованных в одной Х-хромосоме, с диплоидным набором реализаторов мужского П., локализованных в аутосомах, развивается мужской П. И только 2 дозы реализаторов женского П., локализованные в 2 Х-хромосомах, пересиливают потенцию развития мужского П. и тем самым обусловливают женский П. У человека полоопределяющую роль играет Y-хромосома. В аномальных случаях она сочетается с 2, 3 и даже 4 Х-хромосомами при нормальном наборе аутосом. Хотя это и приводит к патологическим отклонениям, однако все особи с такими наборами хромосом бывают мужского П. Полоопределяющая роль Y-хромосом отмечена у многих видов животных, а среди растений - у дрёмы луговой. У дрозофилы Y-хромосома почти не содержит генов, т. е. наследственно инертна; реализаторы женского П. локализованы в Х-хромосоме, реализаторы мужского П. - в аутосомах. Развитие П. контролируется отношением Х-хромосом к набору аутосом (Х: А), условно принятым у самки за единицу (2Х:2А = 1): это отношение у самца равно 0,5 (Х:2А = 0,5). Увеличение этого отношения (полового индекса) свыше единицы приводит к чрезмерному развитию женских половых признаков («сверхсамки»), уменьшение же ниже 0,5 способствует появлению самцов с более выраженными мужскими признаками («сверхсамцы»). Особи с половым индексом 0,67 и 0,75 имеют промежуточное развитие признаков обоих П. и называют интерсексами. Явление
интерсексуальности
демонстрирует бисексуальную потенцию наследственной информации, передаваемой всем потомкам.
Механизм генетического контроля над развитием половых признаков может быть внутри- и межклеточным. Внутриклеточное определение П. не связано с образованием половых гормонов (например, у насекомых), и действие генов, определяющих П., ограничено клетками, в которых эти гены функционируют. При этом в одном организме могут нормально развиваться, не влияя друг на друга, участки тела с женскими и мужскими признаками (см. Гинандроморфизм ) .При межклеточном определении П., характерном для млекопитающих и птиц, под контролем генов вырабатываются половые гормоны,которые, проникая во все клетки организма, обусловливают фенотипическое развитие признаков соответствующего П. Различают прогамное, сингамное и эпигамное определение П. Прогамное определение П. происходит до оплодотворения яйца, например дифференцировка яйцеклеток на быстро и медленно растущие. Первые становятся крупными, и из них после оплодотворения развиваются самки, вторые отличаются меньшими размерами и дают самцов, хотя оба вида яйцеклеток генетически одинаковы. Сингамное определение П. происходит во время оплодотворения, но на разных стадиях этого процесса. У некоторых видов с мужской гетерогаметией и физиологической полиспермией (оплодотворение яйцеклетки несколькими сперматозоидами) П. определяется в момент слияния ядер половых клеток ( кариогамия ) .Если с ядром яйцеклетки сливается мужское ядро с Y-хромосомой, разовьётся мужская особь, если с Х-хромосомой - женская. При женской гетерогаметии П. потомства зависит от того, какая из половых хромосом попадает в ядро яйцеклетки во время мейоза. Если в ядре окажется Z-хромосома, разовьётся особь мужского П., если W-хромосома - женского. Т. о., в данном случае П. зиготы устанавливается до кариогамии. Эпигамное определение П. наблюдается у разнополых видов с фенотипическим определением П., когда направленность развития в сторону мужского или женского П. обусловливается влиянием внешних условий после оплодотворения. Зависимость признаков от пола. Зависят от П. признаки, ограниченные и контролируемые им. Ограниченные П. признаки в силу половой дифференциации могут проявиться только у одного из П. (продукция молока или яиц свойственна только женскому П.), хотя полимерные гены этих признаков локализованы в аутосомах обоих П. Признаки, контролируемые П., проявляются или у обоих П. (с разной степенью выраженности), или (чаще) только у одного из П. (более мощное развитие рогов у баранов, бороды - у козлов), хотя оба в равной мере содержат в аутосомах гены этих признаков. Несходное их развитие обусловлено значительным различием физиологических процессов в организмах разного П. Гены, детерминирующие признаки, сцепленные с П., локализованы как в парных, так и непарных половых хромосомах и поэтому наследуются иначе, чем признаки, обусловленные парными генами, локализованными в аутосомах обоих П. Если гены локализованы в непарной Y-хромосоме гетерогаметного самца, то обусловливаемые ими признаки наследуются лишь сыновьями, а при локализации генов в хромосоме гетерогаметной самки - только дочерьми. Наследуемые т. о. признаки называются голандрическими. Этот тип наследования обнаружен у некоторых видов рыб и насекомых. У др. видов животных он с полной достоверностью не доказан. При локализации генов в гомологичных Х- или Z- хромосомах обусловленные ими признаки передаются сцепленно с П. по типу, получившему название наследования крест-накрест, когда рецессивный признак матери проявится у сыновей, а доминантный - у дочерей (Т. X. Морган), что встречается у многих видов животных (например, трёхцветность кошек, полосатость окраски оперения и скорость его роста у кур). Много сцепленных с П. мутаций обнаружено у дрозофилы и тутового шелкопряда. Сцепленными с П. могут быть и летали - гены, обусловливающие смертельный исход при развитии организма. Если гомогаметный родитель гетерозиготен по летали, локализованной в одной из гомологичных половых хромосом (X или Z), то половина его гетерогаметных потомков погибнет, получив деталь, губительному действию которой в генотипе не будет противопоставлен нормальный аллель.При гетерогаметии женского П. от леталей гибнет половина дочерей, а при гетерогаметии мужского П. - половина сыновей. Иногда мутантные гены в Х- и Z- хромосомах лишь частично снижают жизнеспособность потомства или вызывают различные заболевания, наиболее часто проявляющиеся у гетерогаметного П. У человека обнаружено свыше 50 сцепленных с П. мутаций, приводящих большей частью к нарушению нормальной жизнедеятельности организма (см. Хромосомные болезни ) . Соотношение полов. При фенотипическом определении П. оно зависит от количества развивающихся организмов, которые попадают под влияние внешних факторов, детерминирующих тот или иной П. При генетическом определении П. соотношение П. у большинства видов, как правило, очень близко к 100@: 100B (100 самок: 100 самцов). Однако и при таком определении П. есть отклонения. Так, у некоторых видов млекопитающих с мужской гетерогаметией статистически достоверно рождается на 1-2% больше потомков мужского П. Регуляция пола. Существенный сдвиг соотношения организмов в сторону одного из П. имеет как теоретическое, так и практическое значение, т.к. один из П. обычно более продуктивен. Методы регуляции П., сведённые к 4 основным направлениям, применяются в зависимости от типа определения П. и биологических и хозяйственных особенностей вида. Фенотипическое переопределение П. Если действие генов П. реализуется посредством гормонов, половые признаки изменяются при пересадке половых органов одного П. другому (см. Гонады ) или при введении в организм гормонов противоположного П., а также некоторых аминокислот. Степень фенотипических изменений П. зависит от особенностей вида и дозы введённого препарата. Однако лишь в редких случаях (у некоторых рыб и земноводных) особи с фенотипически переопределённым П. продуцируют гаметы, противоположные их генотипическому П. В следующем поколении, если действие гормонов прекращается, снова вступает в силу генетический механизм определения П. Управление генетическим механизмом определения П., или искусственное сочетание в яйцеклетке половых хромосом. Направленное изменение соотношения П. достигнуто в экспериментах с тутовым шелкопрядом, у которого П. строго определяется сочетанием половых хромосом (ZW - @ ;ZZ - B). Неоплодотворённые яйца после прогрева развиваются партеногенетически (см. Партеногенез ) за счёт диплоидного ядра, не завершившего редукционного деления. Все клетки партеногенетического эмбриона сохраняют материнскую структуру, в частности и в отношении половых хромосом ZW, и, следовательно, развиваются только в самок (Б. Л. Астауров). Воздействием ионизирующих излучений и прогревом удалось подавить в свежеотложенном осеменённом яйце женское ядро и переключить развитие на мужское начало. Диплоидное ядро мужской зиготы образуется путём слияния двух мужских ядер и поэтому имеет структуру мужского П. ZZ. Из таких зигот развиваются гусеницы всегда мужского пола (X. Хасимото; Б. Л. Астауров). Этими методами впервые у с.-х. вида шелкопряда решена проблема произвольной регуляции П. У млекопитающих учёные пытаются разделить по морфологическим и физиологическим особенностям Х- и Y-сперматозоиды с целью последующего осеменения одной категорией сперматозоидов. Однако этим способом пока не удалось достоверно сместить соотношение П. Раннее распознавание П. Используется для сортировки вылупившихся цыплят на петушков и курочек по окраске оперения, сцепленной с П., а также для «сверхранней» сортировки по П. тутового шелкопряда. Под действием ионизирующего облучения у шелкопряда пересажена аутосома с доминантным геном, обусловливающим тёмную окраску яиц тутового шелкопряда, на половую W- хромосому. Сцепление хромосом стойко передаётся по наследству. Те яйца, в которые попадает W- хромосома с пересаженным доминантным геном, приобретают тёмный цвет и развиваются в самок, в то время как яйца мужского П., не получив доминантного гена, остаются непигментированными. Фотоэлектрические автоматы с большой скоростью разделяют разноокрашенные яйца по П. Выведенные таким способом (В. А. Струнников и Л. М. Гуламова) меченые по П. породы шелкопряда находят практическое применение в советском шелководстве. В 60-х гг. 20 в. в опытах английских учёных Р. Эдуардса и Р. Гарднера зафиксировано рождение потомства только одного П. и у млекопитающих. У кроликов извлекали из тела матери ранних зародышей, цитологическим методом определяли их П. и затем зародышей нежелательного П. выбраковывали, а зародышей нужного П. возвращали в матку. Около 20% возвращенных зародышей прижилось и развивалось в крольчат предсказанного учёными П. Изменение соотношения П. может быть почти у всех животных с генетическим определением П. результатом гибели половины зародышей гетерогаметного П. под действием сцепленных с П. деталей. Однако для многих с.-х. животных такой подход к регуляции П. экономически не оправдан. Исключение составляет тутовый шелкопряд. В СССР радиационным методом выведена (В. А. Струнников) генетически особая порода тутового шелкопряда, у которой в обоих Z- хромосомах самцов всегда имеется по одной негомологичной друг другу летали (сбалансированные летали). Если этих самцов скрестить с самками обычных пород, на стадии яйца одна половина самок погибнет от первой, а другая - от второй летали. Из яиц мужского П. вылупляются нормальные гусеницы. Этот способ позволяет в неограниченных количествах получать у тутового шелкопряда только один более продуктивный мужской П. Эволюция пола. Раздельнополость (бисексуальность), свойственная уже многим одноклеточным (водоросли, простейшие), произошла от смешаннополости. Лишь в некоторых случаях (например, при паразитизме ) смешаннополость могла возникнуть вторично из раздельнополости. Так, у паразитических ракообразных наблюдаются все переходы от смешанно- к раздельнополости (например, раздельнополые виды с хорошо развитыми самками и карликовыми самцами - явственный сдвиг в сторону гермафродитизма). Фенотипическое определение П. древнее генетического, т.к. на ранних стадиях эволюции ещё не существовало специального аппарата половых хромосом. Возникающие на определённых этапах эволюции специальные половые хромосомы (рыбы, земноводные) первоначально морфологически неотличимы от аутосом, и о наличии их можно судить только по признакам, сцепленным с П. Вслед за морфологическими различиями между половыми хромосомами и аутосомами возникает дифференцировка между Х- и Y- хромосомой, что делает всё более редкой конъюгацию между ними и затрудняет обмен их участками при кроссинговере (перекресте). Всё это способствует выполнению специфической функции половых хромосом - быть реализаторами женского или мужского П. Полное исчезновение Y- хромосом делает генетическое определение П. ещё более совершенным: П. определяется равновесием между числом аутосом и хромосом. Лит.:Астауров Б. Л., Генетика пола, в сборнике: Актуальные вопросы современной генетики, [М.], 1966; Бреславец Л. П., Определение и наследственность пола у высших растений, «Тр. по прикладной ботанике, генетике и селекции, Сер. 2», 1934, № 6; Рыжков В. Л., Генетика пола, [Хар.], 1936; Лобашев М. Е., Генетика, 2 изд., Л., 1967. В. А. Струнников. 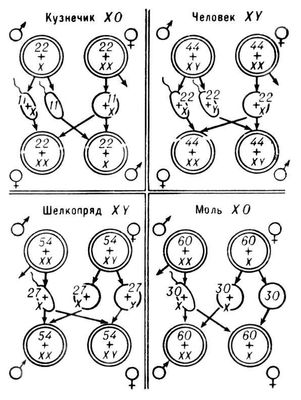
Схема различных типов хромосомного определения пола (двойные кружки - зиготы; одинарные - гаметы; цифры указывают число хромосом). Пол (строит.) Пол, элемент конструкции здания (сооружения), воспринимающий эксплуатационные воздействия от передвижения людей, перемещения грузов, а также от мебели и оборудования, находящихся в помещении. В зависимости от назначения и характера помещения к П. предъявляют определённые требования (конструктивные, эксплуатационные, санитарно-гигиенические, декоративные и др.). П. гражданских зданий должны быть прочными, износостойкими, упругими, гладкими (но не скользкими), обладать малым теплоусвоением, легко очищаться от загрязнения, иметь красивый вид и соответствовать архитектуре интерьера; в мокрых помещениях П., кроме того, должны быть водостойкими и водонепроницаемыми, а в пожароопасных - несгораемыми. В промышленных зданиях к П. предъявляют повышенные требования по сопротивляемости механическим воздействиям (истиранию, удару, продавливанию и др.), а для некоторых производств - по химической стойкости, теплостойкости и др. В современном строительстве конструкция П., как правило, многослойная: П. состоит из основания, в качестве которого могут служить междуэтажные перекрытия здания или грунт, и покрытия (чистого пола) - верхнего лицевого слоя П., непосредственно подвергающегося эксплуатационным воздействиям. Конструкция П. может также включать звуко-, тепло-, гидроизоляционный и др. слои. При устройстве П. по грунту покрытие укладывают на подстилающий слой (подготовку), распределяющий нагрузку по основанию. (В П: по перекрытиям подстилающий слой обычно отсутствует.) Для выравнивания поверхности основания и придания ему необходимой жёсткости поверх вспомогательных слоев обычно делается стяжка (слой цементного раствора), по которой укладывается покрытие. По виду покрытия различают П.: монолитные, или сплошные (цементно-бетонные, асфальтобетонные, мастичные, ксилолитовые, земляные, глинобитные и др.); из листовых и рулонных материалов (линолеум, синтетические ворсовые покрытия, сверхтвёрдые древесноволокнистые плиты и др.); из штучных материалов (паркет, доски, брусчатка, керамическая плитка, клинкерный кирпич, металлические плиты и др.). Наиболее перспективны П. с покрытиями из полимерных рулонных материалов (линолеума, релина и др.), особенно на теплозвукоизоляционной основе, и бесшовные мастичные П. Лит.:Тищенко И. И., Максимова О. М., Устройство полов. Справочник, М., 1972; Белоусов Е. Д., Линде Е. М., Быков А. С., Полы жилых и общественных зданий, М., 1974. К. Н. Попов. Пола Пола', река в Новгородской области РСФСР. Длина 267 км,площадь бассейна 7420 км 2.Берёт начало на северо-западных склонах Валдайской возвышенности, впадает в озеро Ильмень (в низовьях соединяется с р. Ловать). Питание смешанное, с преобладанием снегового. Весеннее половодье, летне-осенняя межень. Средний расход воды 63 м 3/сек.Замерзает в ноябре - декабре, вскрывается в марте - апреле. Сплав леса. Полабские славяне Пола'бские славя'не, полабы, полабяне, большая группа западнославянских племён, населявшая в конце 1 - начале 2-го тыс. н. э. территории от р. Лаба (Эльба) и её притока р. Сала (Заале) на З. до р. Одра (Одер) на В., от Рудных гор на Ю. и до Балтийского моря. на С. Объединялись П. с. в три племенных союза лужицкие сербы, лютичи (велеты), бодричи (ободриты). Основой их хозяйственной деятельности были земледелие и скотоводство, значительного развития достигли садоводство и огородничество. У П. с. интенсивно развивалось ремесло; они вели оживлённую торговлю. С 10 в. германские феодалы начали систематическое наступление на П. с. сначала ради получения дани, а затем - с целью распространения на их землях своей власти путём основания военных областей (марок); католические миссионеры насильственно обращали П. с. в христианство и взимали с них церковную десятину. В 10 в. герм. феодалам удалось подчинить П. с., однако в результате мощных восстаний (983, 1002) большая часть их (за исключением лужицких сербов) вновь стала свободной. Длительные войны с германскими феодалами пагубно сказывались на экономическом развитии П. с., тормозили процесс образования у них относительно крупных раннефеодальных государств. Однако в 10-11 вв. в результате развития классовых отношений процесс этот ускорился. Наиболее значительное государство П. с. - Вендская держава,во главе которого стал князь самой развитой группы П. с. - бодричей - Готшалк.В 12 в. наступил заключительный этап борьбы П. с. во главе с князем бодричей Никлотом против германской агрессии, организаторами которой были Генрих Лев и Альбрехт Медведь. Крестовый поход против славян 1147 окончился безрезультатно для захватчиков. Однако в 50-60-х гг., используя раздробленность П. с. на различные княжения, враждовавшие между собой, германские феодалы захватили последние свободные земли П. с. Земля лютичей стала основой маркграфства Бранденбург,в земле бодричей возникло княжество Мекленбург,вассально зависимое от саксонских герцогов, но управлявшееся славянскими князьями. В 1169 дат. феодалы разрушили святилище П. с. Аркону (на о. Руяна). В последующем большая часть П. с. подверглась германизации, часть была истреблена. Однако некоторая часть П. с. сохранила свою этническую и культурную самобытность (см. Лужичане ) .
Лит.:Die Slawen in Deutschland. Geschichte und Kultur der slawischen Stдmme westlich von Oder und Neisse von 6-12 Jahrhundert. Bearb. von J. Herrmann, 2 Aufl., B., 1972. См. также лит. при статьях Бодричи , Вендская держава , Крестовый поход против славян 1147 . 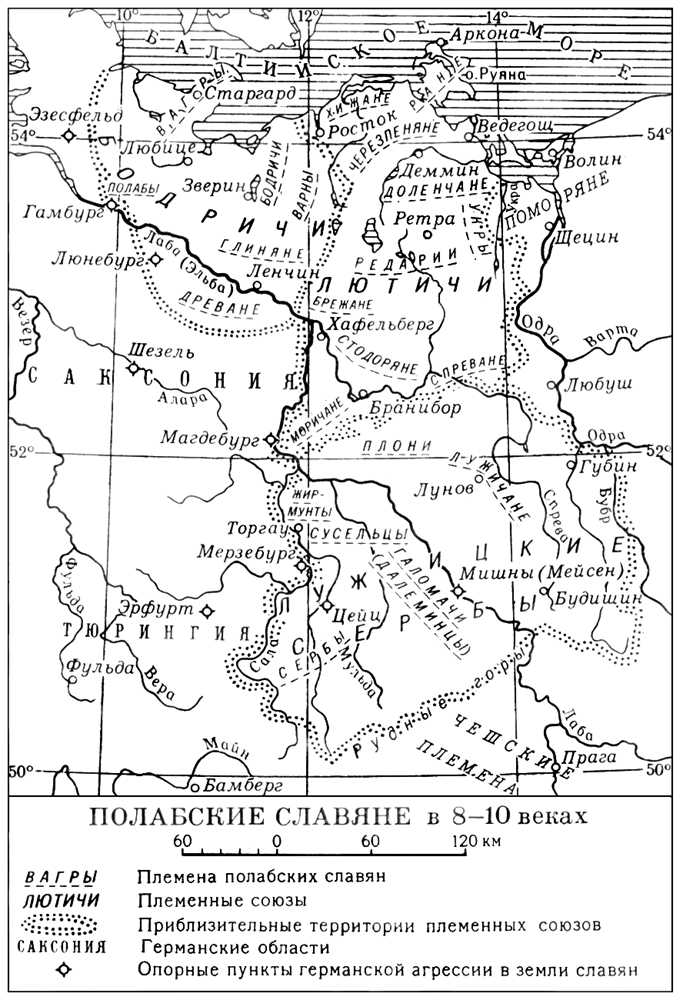
Полабские славяне в 8-10 веках. Полабский язык Пола'бский язы'к, язык полабских славян,живших на территории между нижней Одрой (Одером), нижней и средней Лабой (Эльбой) и Балтийским морем. К концу 17 в. большая часть полабян утратила П. я. Дольше сохранялась славянская речь у древян на крайнем З. (Люховский округ, по р. Этцель), но и там к середине 18 в. вымерла, уступив место немецкому языку. О П. я. можно судить по отдельным словам и фразам, записанным на рубеже 17-18 вв. немецкими учёными (Х. Генниг, И. Парум-Шульце, И. Пфеффингер, Буххольц и др.), по многим следам в топонимике, по отражениям в немецких говорах (лексические «полабизмы», некоторые суффиксы, отсутствие члена, смещение грамматических родов, особенности образования перфекта, утрата начального h- и т.д.). П. я. принадлежит к западной группе славянских языков и обнаруживает преимущественно связи с поморскими и польскими, отчасти лужицкими говорами; подвергся сильному влиянию немецкого языка (обилие дифтонгов, становление артикля, особенности образования сложных времён, перестройка падежной системы, обилие лексических заимствований). Вместе с тем в П. я. сохранялись такие архаизмы, как двойственное число, аорист и имперфект, формы без метатезы (тип tort), некоторые просодические особенности. Лит.:Rost P., Die Sprachreste der Dravдno-Polaben im Hannцverschen, Lpz., 1907; Trubetzkoy N., Polabische Studien, W. - Lpz., 1929; Lehr-SpBawнnski Т., Gramatyka poBabska. Lwуw, 1929; Lehr-SpBawinski Т., PolaDski K., SBownik etymologiczny jzyka Drzewian poBabskich, WrocBaw - Warsz. - Krakow, zesz. 1, 1962; Oles ch R., Fontes linguae Dravaeno-Polabicae minores et chronicavenedica J. P. Schultzii, Kцln - Graz, 1967; его же. Bibliographic zum Dravanopolabischen, Kцln - Graz, 1968. В. Н. Топоров. Полазна Пола'зна, посёлок городского типа в Добрянском районе Пермской области РСФСР. Расположен на берегу Камского водохранилища, в 20 кмк Ю. от г. Добрянка. Добыча нефти и газа. Полак Карл По'лак(Polak) Карл (12.12.1905, Вестерштеде, - 27.10.1963, Берлин), общественный деятель и учёный-юрист ГДР. Родился в крестьянской семье. Ещё в студенческие годы стал марксистом, участвовал в антифашистском движении. В период фашизма эмигрировал в СССР, где вёл научную работу в АН СССР, был секретарём комитета им. Э. Тельмана, боровшегося за освобождение Германии от нацизма. В 1946 вернулся на родину. Внёс существенный вклад в строительство первого социалистического германского государства: был членом Немецкого Народного совета и заместитель председателя его Конституционного комитета, депутат Народной палаты, член Государственного совета, участвовал в научной подготовке важнейших документов СЕПГ, а также мер по совершенствованию государственного органов республики. П. был членом Германской АН в Берлине. Как учёный П. содействовал развитию марксистско-ленинской науки о государстве и праве. Основной труд П. - «Диалектика в учении о государстве» (1959). Награжден «Серебряным орденом за заслуги перед отечеством» и медалью «Борцу против фашизма». Поланецкий универсал 1794 Полане'цкий универса'л 1794, указ, подписанный Т. Костюшко в лагере у местечка Поланец (PoBaniec, близ Сандомежа) 7 мая во время Польского восстания 1794.Составлен при участии Г. Коллонтая.Предусматривал предоставление крестьянам личной свободы при условии их расчёта с помещиками и уплаты государственных налогов, сокращение барщины (особенно на время восстания), признавал за крестьянами наследственное право на обрабатываемую ими землю. П. у., который был шагом вперёд по сравнению с принятой Четырёхлетним сеймом (1788-92) конституцией 1791, практически не был выполнен из-за саботажа шляхты и католического клира. Источн.: Akty powstania Ko[ciuszki, t. 1, Kr., 1918. Полатлы Полатлы'(Polatli), остатки многослойного поселения 3-2-го тыс. до н. э. на окраине города того же названия в Центральной Турции. Исследовались в 1949 (С. Ллойд и Н. Гёкче). В культурном слое (до 24 м) выделен 31 строительный горизонт; они объединены в 4 крупные фазы: 1-я и 2-я относятся к центральноанатолийскому раннему бронзовому веку (середина и 2-я половина 3-го тыс. до н. э.), 3-я - к периоду древнеассирийских колоний в Центральной Анатолии (1-я четверть 2-го тыс. до н. э.), 4-я - к хеттскому периоду (середина 17-12 вв. до н. э.). Для всех фаз характерны прямоугольные дома из камня и сырцового кирпича. Типичная керамика: в 1-й фазе - лепные сосуды с тёмным лощением, во 2-й появляются круговые чаши и сосуды с росписью поверх лощения, в 3-й - гончарный круг и посуда с монохромной росписью, для 4-й фазы характерны узкогорлые кувшины с высоким носиком и фильтром. Металлические изделия и литейные формы встречены во всех фазах. В нижних горизонтах найдены также примитивные глиняные статуэтки. П. - один из основных стратиграфических (см. Стратиграфия ) эталонов культуры бронзового века Центральной Анатолии. Лит.:Lloyd S. and Gцkзe N., Excavations at Polatli, в кн.: Anatolian Studies, v., 1, L., 1951; Orthmann W., Die Keramik der frьhen Bronzezeit aus Inneranatolien, B., 1963. Н. Я. Мерперт. Полба По'лба, полбяная пшеница, группа видов пшеницы с ломким колосом и плёнчатым зерном. При созревании колос распадается на колоски с члениками стержня. Зерно при молотьбе не вымолачивается из плёнок. Виды П.: дикорастущие - дикая двузернянка (Triticum dicoccoides), одноостая однозернянка (Tr. boeticum), двуостая однозернянка (Tr. thaoudar), пшеница Урарту (Tr. urarthu); культурные - двузернянка (Tr. dicoccum), наиболее распространена в культуре, пшеница спельта (Tr. spelta), пшеница маха (Тг. macha), пшеница Тимофеева (Tr. timofeevi). П. отличаются неприхотливостью, скороспелостью, устойчивостью к грибным заболеваниям (большинство видов). В мировом земледелии занимают небольшую площадь. П. - ценный исходный материал для селекции. Полбин Иван Семенович По'лбинИван Семенович [14(27).1.1905, с. Ртищево-Каменка ныне Майнского района Ульяновской области, - 11.2.1945], дважды Герой Советского Союза (23.11.1942 и 6.4.1945), генерал-майор авиации (1943). Член КПСС с 1927. В Советской Армии с 1927. Окончил Оренбургскую военную школу лётчиков (1931). В боях на р. Халхин-Гол командовал бомбардировочным полком. Во время Великой Отечественной войны 1941-45 на различных фронтах, командовал 150-м бомбардировочным авиационным полком (1941-42), 301-й бомбардировочной авиационной дивизией (1942-43), 6-м гвардейским бомбардировочным авиационным корпусом (1943-45). Совершил 157 боевых вылетов на бомбардировку важных военных объектов. Погиб при выполнении боевого задания. Награжден 2 орденами Ленина, 2 орденами Красного Знамени, орденами Суворова 2-й степени, Богдана Хмельницкого, Отечественной войны 1-й степени и медалями. 
И. С. Полбин. Полдень По'лдень, момент, когда для данного места на Земле центр Солнца (истинного или т. н. среднего) находится в верхней кульминации. Прохождению через меридиан истинного Солнца соответствует истинный П., прохождению среднего Солнца - средний П. (см. Время ) .Время наступления П. зависит от географической долготы места: через каждые 15° к З. полдень наступает на 1 час позднее. Полдневица Полдневи'ца, посёлок городского типа в Поназыревском районе Костромской области РСФСР, в 32 кмот ж.-д. станции Супротивный (на линии Буй - Котельнич). Шортюгский леспромхоз. Поле (алгебраич.) По'леалгебраическое, важное алгебраическое понятие, часто используемое как в самой алгебре, так и в др. отделах математики и являющееся предметом самостоятельного изучения. Над обычными числами можно производить четыре арифметических действия (основные - сложение и умножение, и обратные им - вычитание и деление). Этим же характеризуются и П. Полем называется всякая совокупность (или множество) элементов, над которыми можно производить два действия - сложение и умножение, подчиняющиеся обычным законам (аксиомам) арифметики: I. Сложение и умножение коммутативны и ассоциативны, т. е. a+ b= b+ a, ab= ba, a+ ( b+ c) =( a+ b) + c, a( bc) = ( ab) c. II. Существует элемент 0 (нуль), для которого всегда а+ 0 = а;для каждого элемента асуществует противоположный - а,и их сумма равна нулю. Отсюда следует, что в П. выполнима операция вычитания а - b. III. Существует элемент е(единица), для которого всегда ае= а;для каждого отличного от нуля элемента асуществует обратный a -1;их произведение равно единице. Отсюда следует возможность деления на всякое не равное нулю число а. IV. Связь между операциями сложения и умножения даётся дистрибутивным законом: a( b+ c) = ab+ ac. Приведём несколько примеров П.: 1) Совокупность Р всех рациональных чисел. 2) Совокупность Rвсех действительных чисел. 3) Совокупность Квсех комплексных чисел. 4) Множество всех рациональных функций от одного или от нескольких переменных, например с действительными коэффициентами. 5) Множество всех чисел вида
а + b
6) Выбрав простое число р,разобьем целые числа на классы, объединив в один класс все числа, дающие при делении на родин и тот же остаток. Возьмём в двух классах по представителю и сложим их; тот класс, в который попадёт эта сумма, назовем суммой выбранных классов. Аналогично определяется произведение. При таком определении сложения и умножения все классы образуют П.; оно состоит из рэлементов. Из аксиом I, II следует, что элементы П. образуют коммутативную группу относительно сложения, а из аксиом I, III - то, что все отличные от 0 элементы П. образуют коммутативную группу относительно умножения. 1, 2, 3, 4, 5, 6, 7, 8, 9, 10, 11, 12, 13, 14, 15, 16, 17, 18, 19, 20, 21, 22, 23, 24, 25, 26, 27, 28, 29, 30, 31, 32, 33, 34, 35, 36, 37, 38, 39, 40, 41, 42, 43, 44, 45, 46, 47, 48, 49, 50, 51, 52, 53, 54, 55, 56, 57, 58, 59, 60, 61, 62, 63, 64, 65, 66, 67, 68, 69, 70, 71, 72, 73, 74, 75, 76, 77, 78, 79, 80, 81, 82, 83, 84, 85, 86, 87, 88, 89, 90, 91, 92, 93, 94, 95, 96, 97, 98, 99, 100, 101, 102, 103, 104, 105, 106, 107, 108, 109, 110, 111, 112, 113, 114, 115, 116, 117, 118, 119, 120, 121, 122, 123, 124, 125, 126, 127 |
|||||||
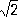 ,где
аи
b- рациональные числа.
,где
аи
b- рациональные числа.